

Abstract
Neutrophils act as the body’s first line of defense against infection and respond to diverse inflammatory cues, including cancer. Neutrophils display plasticity, with the ability to adapt their function in different inflammatory contexts. In the tumor microenvironment, neutrophils have varied functions and have been classified using different terms, including N1/N2 neutrophils, tumor-associated neutrophils, and polymorphonuclear neutrophil myeloid–derived suppressor cells (PMN-MDSCs). These populations of neutrophils are primarily defined by their functional phenotype, because few specific cell surface markers have been identified. In this review, we will discuss neutrophil polarization and plasticity and the function of proinflammatory/anti-inflammatory and protumor/antitumor neutrophils in the tumor microenvironment. We will also discuss how neutrophils with the ability to suppress T-cell activation, referred to by some as PMN-MDSCs, fit into this paradigm.
Cancer as an unhealed wound
Neutrophils are the first responders to tissue damage and play a critical role in host defense against infection. However, persistent neutrophil infiltration is a hallmark of chronic inflammation and contributes to tissue damage. The tumor microenvironment is characterized by this type of chronic inflammation and has been described as the wound that does not heal. This unresolved tissue inflammation can mediate tumorigenesis, as has been described in inflammatory bowel disease, increasing the risk of colon cancer, and nonalcoholic fatty liver disease or chronic hepatitis, leading to progression to hepatocellular carcinoma.1-3 In addition to having tumor-promoting roles in the context of innate immune inflammation and tumor initiation, neutrophils can also promote tumor progression by suppressing the function of the adaptive immune response in the tumor microenvironment. There is growing interest in targeting this type of suppressive function, referred to as myeloid-derived suppressor cell function, to optimize for T-cell antitumor activity and immunotherapy efficacy against cancer.
Neutrophils in wounds and the cancer microenvironment
Neutrophils are the most prevalent type of innate immune cell and are the first cells to arrive at sites of developing inflammation. The dynamic behavior of neutrophils at wounds has been demonstrated in model organisms, like zebrafish, where the temporal and spatial behaviors of neutrophils can be visualized. In these models, neutrophils are often the first cells to arrive both at a wound and during the early initiation phases of carcinogenesis. For example, in zebrafish models of cancer where early tumor initiation can be imaged, expression of oncogenic RasG12V in zebrafish skin melanocytes or epidermal cells induces early recruitment of neutrophils, which drive cell proliferation4 and epithelial to mesenchymal transition.5 The early tissue cues that recruit neutrophils to wounds and transformed cells have common mechanisms, including the involvement of damage-associated molecular patterns and specific chemokines.6,7 One of the first chemokines produced in both wounds and cancer is interleukin-8 (IL-8). It has been shown in the zebrafish model that IL-8 recruits neutrophils via the CXCR1/CXCR2 receptors to both tissue damage and cancer.8,9 Furthermore, in some breast cancer models, inhibiting CXCR2 reduces neutrophil recruitment to tumors and increases the efficacy of chemotherapy.10 These results suggest that neutrophil recruitment to cancer can be targeted through pathways that also mediate recruitment to wounds, with a beneficial effect on patient outcome. Indeed, neutrophils have been implicated in metastasis through both leukotriene-generating enzyme arachidonate 5-lipoxytenase (Alox5)11 and neutrophil-generated transferrin.12 However, some neutrophils may have antitumor effects in cancer, and the ability to specifically target the protumor neutrophils while preserving the function of antitumor neutrophils is a future challenge.
Inflammation resolution and neutrophil reverse migration
Neutrophils are much longer lived than initially suggested, and growing evidence supports their plasticity and ability to adapt their phenotype depending on the tissue environment.13-16 After acute inflammation, neutrophil resolution is critical to prevent tissue damage and transition to chronic wounds or damage-induced cancer. Neutrophil resolution can occur through neutrophil apoptosis and subsequent clearance by macrophages. Recent studies show, through direct visualization, neutrophils exit damaged tissues and reverse migrate back into the bloodstream. This reverse migration and reverse transendothelial migration, first observed in zebrafish17 and more recently in mice,18,19 suggest that neutrophil function at sites of tissue damage and cancer may be more complex than initially understood. The recent finding that neutrophils leave sites of sterile inflammation in the liver and traffic to the lung and bone marrow in mice raises intriguing questions about the function of reverse migration in cancer.18 Because the lung and bone are common sites of metastatic disease, it is intriguing to speculate that neutrophils could be promoting metastasis to specific niches through neutrophil reverse migration. However, the role of neutrophil recruitment and resolution in the context of cancer still remains mostly unknown.
Neutrophil subtypes and plasticity in inflammation
Because of the variety of inflammatory stimuli to which neutrophils respond, they must be able to adapt to different environments. Therefore, it is not surprising that neutrophils with varying phenotypes have been described in different inflammatory contexts. The classification of neutrophil populations is a controversial topic. Some studies suggest that neutrophil populations should be regarded as subgroups of neutrophils, but many of these subgroups lack distinctive markers, especially in humans, and are instead defined by their phenotypes. An example of changing neutrophil phenotype is perhaps best characterized by immature and aged neutrophils where cell surface markers have been identified. In humans, immature neutrophils are CD16lo, whereas aged neutrophils can be identified by the surface expression of Cxcr4hi and CD62lo.20 However, these surface changes occur with activation or aging rather than as representation of a developmental program, such as occurs with Th1/Th2 lymphocytes.
It is possible that the necessity for neutrophil recruitment in a wide variety of contexts requires them to maintain phenotypic plasticity. This would suggest that neutrophils with varied functions are adapting to their environment and becoming differentially polarized rather than starting as a specific subtype. This idea is supported by the presence of specific surface markers on reverse-migrated neutrophils.21 A distinct subpopulation of reverse-migrated neutrophils has been identified by CD54hi and Cxcr1lo, and this population is increased in the circulation of patients with inflammation, including acute pancreatitis.21 Another example is the recent study of neutrophil–dendritic cell hybrids,22-24 which demonstrated these can play a role in antigen presentation and potentially have antitumor effects. These cells retain cell surface markers and functional capabilities of both cell types and thus represent an additional example of the plasticity of neutrophils.
The scientific literature focusing on neutrophils in cancer is complicated by the identification of different subsets with unclear markers and with potentially overlapping functions. As a broad initial classification and for the purpose of clarity in understanding neutrophil function in cancer, we will discuss neutrophils as being either proinflammatory or antiinflammatory and the implications of these functions in cancer progression.
Neutrophil polarization states in cancer
Neutrophil polarization may influence the role they play in the tumor microenvironment. It has been shown through meta-analysis that a high neutrophil/lymphocyte ratio in solid tumors is correlated with poor patient outcome.25 However, these data alone do not elucidate whether neutrophils are responsible for cancer progression or merely correlate with advanced disease. In fact, in murine cancer models, the effect of neutrophil infiltration on cancer development has been highly variable. In 2009, Fridlender et al26 first suggested a delineation between antitumorigenic and protumorigenic neutrophils, termed N1 and N2, respectively. They showed that transforming growth factor-β (TGF-β), an immunosuppressive cytokine overexpressed by tumor cells, polarized neutrophils to a protumorigenic phenotype (N2) and that neutrophil depletion caused a small decrease in tumor growth in mouse models. However, TGF-β blockade, using the TGF-β receptor inhibitor SM16, led to an accumulation of neutrophils with an antitumorigenic phenotype (N1). In this case, neutrophil depletion led to increased tumor growth. In addition to increasing after TGF-β blockade, N1 neutrophil polarization is thought to be induced by type 1 interferons (IFNs).27,28 Therefore, the phenotypic switch from N1 to N2 may suggest antagonistic signaling pathways between TGF-β and type 1 IFN cytokines.27
The N1/N2 naming scheme was modeled after the activation states seen in tumor-associated macrophages. Despite functional differences, no definitive surface marker has been identified to distinguish N1 and N2 tumor-associated neutrophils (TANs),26 indicating these subpopulations may represent neutrophils with differential activation states. Overall, these results indicate that neutrophil polarization is likely driven by the tumor microenvironment. It was recently shown that neutrophils from mice with early-stage tumors had a higher level of spontaneous migration compared with neutrophils from late-stage tumors, indicating a phenotypic switch mediated by the tumor microenvironment.29 Furthermore, neutrophils themselves can influence the tumor microenvironment, both directly and indirectly through other cell types. We will first describe their direct effects and then delve further into understanding how neutrophils can shape the tumor landscape through pro- or anti-inflammatory signals. A schematic depicting the many roles neutrophils can play in the cancer microenvironment is provided in Figure 1. Because of the difficulty of manipulating primary neutrophils, much of the published literature on TANs refers to studies done in mice. However, we will focus primarily on studies using human neutrophils in an effort to highlight the need for further progress in this area.
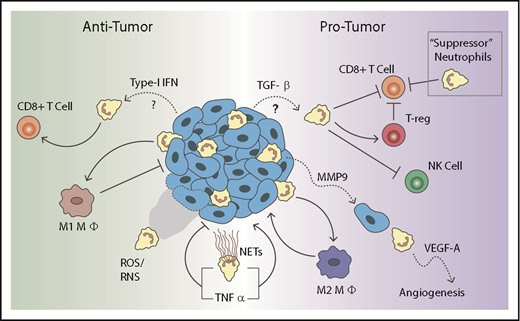
Neutrophils in the tumor microenvironment. Neutrophils can either promote or inhibit tumor growth depending on their polarization states. Antitumor neutrophils can directly kill tumor cells through release of reactive oxygen species (ROS) and reactive nitrogen species (RNS). They can also promote T-cell activation and recruit proinflammatory (M1) macrophages. In contrast, protumor neutrophils can release matrix metalloproteinase 9 (MMP9), which promotes angiogenesis and dissemination of tumor cells. They can also suppress natural killer (NK) cell function. Furthermore, they can recruit anti-inflammatory (M2) macrophages and T-regulatory cells. Finally, suppressor neutrophils, often referred to as polymorphonuclear neutrophil myeloid-derived suppressor cells (PMN-MDSCs), as well as other protumor neutrophils, can suppress CD8 T-cell function.
Neutrophils in the tumor microenvironment. Neutrophils can either promote or inhibit tumor growth depending on their polarization states. Antitumor neutrophils can directly kill tumor cells through release of reactive oxygen species (ROS) and reactive nitrogen species (RNS). They can also promote T-cell activation and recruit proinflammatory (M1) macrophages. In contrast, protumor neutrophils can release matrix metalloproteinase 9 (MMP9), which promotes angiogenesis and dissemination of tumor cells. They can also suppress natural killer (NK) cell function. Furthermore, they can recruit anti-inflammatory (M2) macrophages and T-regulatory cells. Finally, suppressor neutrophils, often referred to as polymorphonuclear neutrophil myeloid-derived suppressor cells (PMN-MDSCs), as well as other protumor neutrophils, can suppress CD8 T-cell function.
Direct effects
Neutrophils are capable of both killing tumor cells and promoting tumor growth in differing contexts. The most well-known example of neutrophil-directed cytotoxicity against tumor cells is ROS production (superoxide and hydrogen peroxide), but the mechanism of and role played by ROS in the tumor microenvironment are unclear. Recently, ROS-mediated cell killing was shown to be dependent on tumor cell expression of TRPM2, an H2O2-dependent Ca2+ channel, which upon activation results in a lethal influx of calcium ions into the cell.30 Furthermore, TANs can also inhibit metastatic seeding in the lungs through hydrogen peroxide generation.31 In addition to ROS, neutrophils secrete RNS, such as nitric oxide and peroxynitrite, but the effect on human cancer cells is not well studied. One mechanism of RNS-induced cancer cell killing was described by Finisguerra et al,32 who found that MET receptor signaling in neutrophils after hepatocyte growth factor binding caused increased nitric oxide production and promoted cancer cell killing.
Neutrophils are also capable of directly promoting tumor progression, metastasis, and angiogenesis. In addition to their cytotoxic roles, ROS and RNS production can be protumorigenic, contributing to oxidative DNA damage and genetic instability.33 Furthermore, release of neutrophil-generated enzymes can promote tumor progression. Mature neutrophils possess granules containing a reservoir of enzymes, including myeloperoxidase (MPO), neutrophil elastase (NE), and MMPs: neutrophil collagenase (MMP8) and gelatinase B (MMP9). MMPs are capable of remodeling the extracellular matrix to promote angiogenesis34 and were recently shown to promote cancer migration and invasion by stabilizing integrins.35 The proangiogenic activity of neutrophils may also be induced by modifications in cell signaling pathways. A recent publication correlated increased nicotinamide phosphoribosyltransferase signaling with a protumor neutrophil phenotype and found increased expression of nicotinamide phosphoribosyltransferase in neutrophils from cancer patient samples.36 In addition to MMPs, other neutrophil enzymes can promote tumor growth. NE has been shown to promote tumor cell proliferation through increased signaling between phosphatidylinositol 3-kinase and the potent mitogen platelet-derived growth factor receptor.37 Additionally, it is known that NE and MPO regulate the production of NETs. Neutrophils typically produce NETs during infection, but when they are released in the tumor microenvironment, they can stimulate migration and invasion of cancer cells.38
In addition to directly affecting the tumor microenvironment, neutrophils are capable of influencing other cell types to promote or inhibit tumor growth and progression. However, the separation of these functions as being strictly immunosuppressive or proinflammatory is an oversimplification, because they can have variable impacts on the tumor microenvironment. A summary of the literature describing the effects of neutrophils in the tumor microenvironment is provided in Table 1.
Human TAN and PMN-MDSC protumor and antitumor functions
| TANs/PMN-MDSCs | Reference | T-cell suppression confirmed in humans | ||
|---|---|---|---|---|
| TANs | PMN-MDSCs | |||
| Protumor | ||||
| T/NK cell suppression | ||||
| Arginase 1 | TANs, PMN-MDSCs | 59 | 74,82,-84,91 | Yes |
| ROS | TANs, PMN-MDSCs | 92 | 84,85 | No |
| STAT3 | PMN-MDSCs | 93 | Yes | |
| STAT/IRF-8 | PMN-MDSCs | 78 | No | |
| LOX-1 | PMN-MDSCs | 89,90 | Yes | |
| mPR3 | TANs | 61 | N/A | |
| MPO/hydrogen peroxide | TANs | 62,63 | N/A | |
| Angiogenesis/metastasis | ||||
| VEGF | TANs, PMN-MDSCs | 94 | 83 | Yes |
| MMP9 | TANs, PMN-MDSCs | 34,95 | 88 | Yes |
| NAMPT/STAT3 | TANs | 36 | N/A | |
| Oncostatin M | TANs | 45,46 | N/A | |
| NETs | TANs | 38 | N/A | |
| TNF-α | TANs | 55 | N/A | |
| IL-8 | TANs | 43,44,59 | N/A | |
| Elastase | TANs | 37 | N/A | |
| Immune cell recruitment | ||||
| CCL17 | TANs | 39,40 | N/A | |
| CCL2 | TANs | 39 | N/A | |
| Antitumor | ||||
| Tumor cytotoxicity | ||||
| ROS | TAN | 30,96 | N/A | |
| Granzyme B | TAN | 97 | N/A | |
| MET signaling | TAN | 32 | N/A | |
| Trogoptosis | TAN | 52 | N/A | |
| TNF-α | TAN | 32,53 | N/A | |
| T-cell activation/priming | ||||
| Mechanism unknown | TAN | 48,98 | N/A | |
| NETs | TAN | 49 | N/A | |
| TANs/PMN-MDSCs | Reference | T-cell suppression confirmed in humans | ||
|---|---|---|---|---|
| TANs | PMN-MDSCs | |||
| Protumor | ||||
| T/NK cell suppression | ||||
| Arginase 1 | TANs, PMN-MDSCs | 59 | 74,82,-84,91 | Yes |
| ROS | TANs, PMN-MDSCs | 92 | 84,85 | No |
| STAT3 | PMN-MDSCs | 93 | Yes | |
| STAT/IRF-8 | PMN-MDSCs | 78 | No | |
| LOX-1 | PMN-MDSCs | 89,90 | Yes | |
| mPR3 | TANs | 61 | N/A | |
| MPO/hydrogen peroxide | TANs | 62,63 | N/A | |
| Angiogenesis/metastasis | ||||
| VEGF | TANs, PMN-MDSCs | 94 | 83 | Yes |
| MMP9 | TANs, PMN-MDSCs | 34,95 | 88 | Yes |
| NAMPT/STAT3 | TANs | 36 | N/A | |
| Oncostatin M | TANs | 45,46 | N/A | |
| NETs | TANs | 38 | N/A | |
| TNF-α | TANs | 55 | N/A | |
| IL-8 | TANs | 43,44,59 | N/A | |
| Elastase | TANs | 37 | N/A | |
| Immune cell recruitment | ||||
| CCL17 | TANs | 39,40 | N/A | |
| CCL2 | TANs | 39 | N/A | |
| Antitumor | ||||
| Tumor cytotoxicity | ||||
| ROS | TAN | 30,96 | N/A | |
| Granzyme B | TAN | 97 | N/A | |
| MET signaling | TAN | 32 | N/A | |
| Trogoptosis | TAN | 52 | N/A | |
| TNF-α | TAN | 32,53 | N/A | |
| T-cell activation/priming | ||||
| Mechanism unknown | TAN | 48,98 | N/A | |
| NETs | TAN | 49 | N/A | |
LOX-1, low-density lipoprotein receptor-1; mPR3, membrane-associated proteinase 3; N/A, not applicable; NAMPT, nicotinamide phosphoribosyltransferase; NET, neutrophil extracellular trap; STAT3, signal transducer and activator of transcription 3; TNF-α, tumor necrosis factor α; VEGF, vascular endothelial growth factor.
Proinflammatory protumor
Proinflammatory neutrophils recruit other immune cells that can have differential effects on the tumor microenvironment, including cells that promote tumor growth. Proinflammatory neutrophils in the tumor microenvironment secrete chemokines, such as CCL2 (monocyte chemoattractant protein-1) and CCL17, which recruit monocytes and T regulatory cells, respectively.39,40 T regulatory cells are known to suppress other inflammatory T-cell populations, leading to increased tumor growth. The role of monocyte recruitment in tumor progression is less clear, because monocyte recruitment can induce or inhibit tumor growth, depending on the polarization state in the tumor microenvironment.41,42
Secretion of cytokines and growth factors by TANs can also promote tumor cell extravasation and metastasis. IL-8 secretion causes more neutrophils to be recruited to the tumor site and leads to PMN confinement and immobilization in the tumor. This localized secretion of IL-8 by confined neutrophils can then promote tumor cell extravasation as a result of increased intracellular adhesion molecule 1 interactions with neutrophils.43,44 Oncostatin M, a member of the IL-6 family, is secreted by neutrophils and can have a variety of proinflammatory and angiogenic effects, including inducing neutrophil adhesion and chemotaxis, increasing chemokine production by endothelial cells,45 and stimulating vascular endothelial growth factor production.46 Finally, TGF-β, produced by hepatic stellate cells in hepatocellular carcinoma, has been shown to promote recruitment of protumorigenic neutrophils. However, depletion of these neutrophils resulted in a decrease in TGF-β–expressing hepatic stellate cells, indicating a possible feedback loop for TGF-β.47
Proinflammatory antitumor
Proinflammatory neutrophils can also inhibit tumor growth by orchestrating the recruitment and function of other immune cells in the tumor microenvironment. In particular, neutrophils are capable of presenting antigens to T cells to stimulate an adaptive immune response and IFN-γ production.48 Neutrophils also interact with T cells via NETs. As previously described, NETs can promote cancer metastasis, but they can also directly prime T cells by reducing their activation threshold.49 Finally, it has been shown that IFN-γ–stimulated neutrophils recruit and further activate NK cells via secretion of IL-18.50
In addition to inhibiting tumor growth through recruitment of other immune cells, neutrophils are also capable of mediating antibody-dependent cellular cytotoxicity51 ; however, the exact mechanism by which neutrophils kill antibody-opsonized cancer cells has only recently been elucidated. Matlung et al52 have proposed that neutrophil antibody-dependent cellular cytotoxicity utilizes antibody-mediated trogoptosis, which involves neutrophil endocytosis and direct disruption of the cancer cell plasma membrane.
Finally, inflammatory molecule secretion by neutrophils can also reduce tumor burden. In 2009, Fridlender et al identified an increased expression of TNFα in mouse N1 cells.26 This expression was shown to promote tumor cell death by priming neutrophils for superoxide release.53,54 Additionally, TNFα, although not alone, was capable of upregulating MET expression and promoting antitumor activity of neutrophils in a variety of cancer types.32 However, the role of this cytokine in the tumor microenvironment is not clear, because TNFα was also found to stimulate melanoma extravasation and metastasis.55 Follow-up analysis of N1 RNA expression profiles in mice indicated that TANs do not change expression of TNFα during tumor progression. Therefore, the role of this cytokine in different neutrophil polarization states requires further study.56
Anti-inflammatory antitumor
Few examples of neutrophil immunosuppressive antitumor behavior exist in the literature, which may stem from little research in this field rather than a lack of these functions. There is some evidence that neutrophil modulation of IL-17 secretion can reduce tumor growth. Neutrophils induce oxidative stress via ROS to suppress IL-17–producing γδ T cells. These cells promote tumor growth and metastasis by secreting IL-17, and neutrophil depletion increased their accumulation in a mouse model of melanoma.57 Another recent study found that IL-17-γδ T cells can polarize neutrophils to an immunosuppressive phenotype.58 Therefore, the interactions between neutrophils and these T cells in the tumor microenvironment are complex and warrant further investigation.
Anti-inflammatory protumor
Neutrophils can have an immunosuppressive role in cancer that promotes tumor growth, primarily by dampening the recruitment of other immune cells to the tumor microenvironment. In particular, neutrophils can release ROS,33 as well as enzymes such as arginase 1, that inhibit the T-cell response to the tumor microenvironment. Intriguingly, IL-8 secretion from cancer cells can induce neutrophils to release arginase into the tumor microenvironment.59 This enzyme degrades extracellular arginine, a protein essential for T-cell activation and proliferation,60 and therefore inhibits the T-cell response. Membrane-associated proteinase 3 is a serine protease, and its enzymatic activity on PMNs was shown to inhibit T-cell proliferation in vitro.61 Neutrophils can also suppress NK cell activity through release of MPO and hydrogen peroxide,62,63 as well as through expression of CXCR4 on their membrane, which correlated with decreased secretion of IL-18, the cytokine responsible for NK cell activation.64 Finally, neutrophils can induce apoptosis of CD-8+ T cells in the tumor environment, via a TNFα and nitric oxide contact–dependent mechanism.65
PMN-MDSCs
A second classification of protumor antiinflammatory neutrophils was identified at approximately the same time as the N1/N2 classification. MDSCs were first described in 200766 as a population of myeloid cells enriched in cancer patients with immunosuppressive function.67 Additional studies have determined that MDSCs are composed of 2 groups: PMN-MDSCs and monocytic MDSCs (M-MDSCs). These 2 cell types share phenotypic and morphologic features and are characterized by 2 specific traits: an immature myeloid state and immunosuppressive properties. Different theories exist around the origin of PMN-MDSCs: acquisition of immunosuppressive features from inflammatory stimuli, immature cells prematurely leaving the bone marrow, modification in granulopoiesis in the bone marrow, or extramedullary granulopoiesis.16 A recent study demonstrated induction of human PMN-MDSCs by unstimulated CD4+ T cells that required cell-cell contact and TNFα secretion. However, surface marker expression (CD33+CD11b+CD16+CD14−) was the only evidence that they were MDSCs, because T-cell suppression was not tested.68 The hallmark of MDSCs is their ability to suppress T cells, but research has provided evidence for modification of NK cell activity and innate immune response as well. However, PMN-MDSCs have not been well studied, particularly in humans, and characterization of their protumorigenic functions is complicated by their very recent distinction from M-MDSCs and limited markers. Here we describe what is known about PMN-MDSCs and raise the question as to the appropriate nomenclature for this population in humans.
PMN-MDSC isolation
Efforts to isolate PMN-MDSCs have been hampered by their lack of specific surface markers in humans. Initial work on characterizing MDSCs in mice used expression of the surface marker CD11b+GR-1+.67 However, the GR-1 surface antigen cannot differentiate between M-MDSCs and PMN-MDSCs. Furthermore, it is only present in mice, and therefore, a different identification scheme is required in human PMN-MDSCs (CD11b+CD14−CD15+/CD66b+). With this marker set, there is still some difficulty in distinguishing between PMN-MDSCs and neutrophils. CD11b and CD33 are nonspecific and expressed on NK cells and all cells in the myelocytic lineage.69,70 CD14 and CD15 are often included, but CD15 is also expressed on eosinophils and mature neutrophils.71,72 CD16 has been proposed as an additional marker because it increases expression with maturation; however, it is not sufficient alone, because it is also expressed on NK cells and monocytes.16 Therefore, defining MDSCs as a strictly immature population may not be correct. An attempt to standardize MDSC characterization in humans involved analysis of the same healthy peripheral blood sample among a consortium of laboratories. Interlaboratory variability was high,73 demonstrating the difficulty of MDSC identification and the need for further study of surface markers to distinguish between cell subsets. Recently, a new marker, CD13+, was identified on MDSCs in human patients with pancreatic ductal adenocarcinoma. CD13hi PMN-MDSCs were more immunosuppressive than CD13lo MDSCs and were correlated with poor prognosis.74
A second method for MDSC isolation utilizes density centrifugation to separate a less granular population of PMN-MDSCs present in the mononuclear layer from the high-density neutrophils (HDNs) in the granulocyte layer.75 These low-density neutrophils (LDNs) are capable of suppressing CD8 T-cell proliferation and comprise a small proportion of the peripheral blood of healthy individuals but are increased in cancer patients. Of note, the LDN population was found to be a heterogeneous mix of both mature and immature cells, identified by segmented and banded neutrophils in cytospin images. Furthermore, HDNs and LDNs are capable of switching phenotypes. Specifically, TGF-β can cause a transition from HDN to LDN and induce immunosuppressive properties.75 This study identified a neutrophil phenotype capable of a specialized immunosuppressive function, but because of a mixed population, the authors could not definitively say that the only cells responsible for this phenotype were immature. Isolation by density centrifugation is also limited, because it can only isolate MDSCs in peripheral blood samples and cannot be used to separate cells from tumor tissues.
Evaluation of neutrophil nuclear morphology is a classic method for determining neutrophil maturation stage. The nucleus in early progenitors is a horseshoe shape, termed banded. The nuclear shape then becomes segmented with maturation. PMN-MDSCs are often described as having a banded nucleus, or a ring-like structure in mice.76 Similarly, Fridlender et al26 characterized N1 neutrophils as having a hypersegmented structure compared with N2 neutrophils. Perhaps this is another example of PMN-MDSC and TAN similarity. However, many other publications have indicated no difference, and this question of nuclear morphology phenotypes has been more extensively reported in other reviews.16
Both PMN-MDSC isolation methods generate heterogeneous populations of cells and demonstrate that the capability to suppress T cells is not restricted to an immature population. Therefore, characterization of MDSCs as a distinct population of immature cells may be too restrictive. Additionally, the influence of TGF-β on HDN-to-LDN switching demonstrates their inherent plasticity and also raises questions about the relationship between TANs and PMN-MDSCs. Although surface marker expression still requires progress for better separation methods, the true test of PMN-MDSC function has been immunosuppressive effects. We will discuss the available literature on this topic in an effort to determine where the functional characterization of PMN-MDSCs and TANs overlap and if in fact this terminology is appropriate based on current knowledge.
Characterization of PMN-MDSCs
Many studies have attempted to further characterize MDSC development by understanding their transcriptional regulation in both mice and humans.77 In particular, granulocyte-colony stimulating factor (G-CSF) and granulocyte-macrophage CSF are implicated in MDSC regulation via the STAT3 and STAT5 pathways.78 In vitro stimulation of CD33+ peripheral blood mononuclear cells with G-CSF was capable of generating a PMN-MDSC population with T-cell immunosuppressive function.68 G-CSF is known to be an important regulator of neutrophils, and elevated levels can promote hyperproliferation and release of immature neutrophils from the bone marrow.79 Furthermore, G-CSF is highly expressed by many tumors and could potentially induce an immunosuppressive phenotype in the tumor microenvironment.80 IRF-8, a transcription factor involved in myeloid differentiation and lineage commitment, is also regulated via the STAT3/5 pathways. This transcription factor was found to be downregulated in patient neutrophils and inversely correlated with MDSC frequency.78 Other work in the field argues that a single signal, such as G-CSF, may not be enough to drive MDSC expansion, and a second inflammatory signal (ie, IL-6) may be required for MDSC activation.81
Study of PMN-MDSCs in humans is limited. Arginase 1 expression has been the most emphasized feature of MDSCs, because it directly inhibits T-cell proliferation and activation. Studies of arginase in human PMN-MDSCs have identified increased expression levels in the blood of multiple cancers and shown correlation with suppressive T-cell function ex vivo.82-84 A second hallmark of MDSC function is the production of ROS. PMN-MDSCs from the peripheral blood of patients with head and neck cancer or non–small-cell lung cancer had higher levels of ROS production upon stimulation with phorbol myristate acetate.84,85 Unlike TANs and M-MDSCs, PMN-MDSCs do not seem to generate nitric oxide.86 S100A8/9 calcium binding proteins, which promote PMN-MDSC migration into tumors in mouse models, were increased in the serum of patients with glioblastoma.82 However, the chemotactic effect of S100A8/9 proteins on PMN-MDSCs in humans is not clear. Additionally, PMN-MDSCs from the peripheral blood of patients display impaired chemotactic activity, which could limit their migration to tumor sites.29,87 Finally, renal cell carcinoma patient MDSCs had high levels of MMP8 and MMP988 and vascular endothelial growth factor.83 Many of these same immunosuppressive and proangiogenic factors are highly expressed by TANs.
Transcriptomic analysis
Transcriptomic analysis has been used to try to distinguish these neutrophil populations. A transcriptomic study evaluated PMNs from healthy individuals and cancer patients, as well as PMN-MDSCs from cancer patients alone. No tumor samples were taken, because these 2 cell types cannot be distinguished in tissues through surface marker expression. Instead, PMN/PMN-MDSCs were isolated from the peripheral blood by density centrifugation (HDN/LDN) and further purified with CD15 bead selection. This study is unique because it allowed analysis of 2 samples from the same patient. PMN-MDSCs from cancer patients had a more distinct gene expression profile than PMNs from the same patient or other healthy individuals.89 For example, the endoplasmic reticulum stress response was elevated, along with high expression of LOX-1, which has not previously been studied in neutrophils. LOX-1+ PMN gene expression profiles highly correlated with PMN-MDSCs and displayed increased T-cell suppression.89 Furthermore, a later study found that LOX-1+CD15+ PMN-MDSCs are increased in patients with hepatocellular carcinoma, and these cells are capable of suppressing T-cell proliferation and IFN-γ production and produce elevated levels of arginase and ROS.90 Additionally, staining for LOX-1+ PMN-MDSCs in the tissue correlated with peripheral blood levels,90 but it would be important to functionally compare LOX-1+ and LOX-1− TANs within tissue samples. Condamine et al89 argued that LOX-1 is not only associated with MDSCs but also critical to their phenotype. However, the study does not address if LOX-1 is responsible for increased immunosuppressive activity. LOX-1 may be a promising candidate, but it was only expressed on one-third of PMN-MDSCs and does not exist in mice.89
More work needs to be done to determine if PMN-MDSCs represent a unique subpopulation of neutrophils or simply represent a continuum of functional states. Further support for human PMN-MDSCs representing an immunosuppressive polarization state is provided by the finding that stimulation of human peripheral blood neutrophils in vitro was sufficient to induce MDSC-like activity, whereas resting neutrophils were not capable of suppressing T-cell proliferation.15 Until surface markers can effectively distinguish PMN-MDSCs, TANs, and neutrophils both in blood and in tumor tissue, it may be best to define these cells as neutrophils with distinct functions or phenotypes. In fact, many papers describing the study of TANs or PMN-MDSCs use the same isolation strategies, further raising the question about the use of the specific term PMN-MDSC. Perhaps human PMN-MDSCs should instead be referred to as neutrophils that suppress T-cell activation until there are better surface markers of these populations.
In conclusion, recent advances highlight the complexity and importance of understanding neutrophil biology in the tumor microenvironment. It is clear that neutrophils can provide both protumor and antitumor functions through the differential regulation of components of the tissue microenvironment and other immune cells. However, a gap remains in understanding how these different neutrophil populations are related and if in fact they represent subtypes or represent dynamic and plastic changes in the phenotype of neutrophils over time. The identification of surface markers enabling the tracking of neutrophils temporally and spatially within the tumor microenvironment will be required for increased understanding of these different populations in humans. The possibility that these phenotypes could be targeted for therapeutic advantage is the challenge of future investigation and may be critical for the successful application of immunotherapy for many patients with cancer.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgment
This work was supported by National Institutes of Health, National Institute of General Medical Sciences grant R35 GM1 18027 01.
Authorship
Contribution: M.A.G., L.E.H., and A.H. wrote and edited the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Anna Huttenlocher, 1550 Linden Dr, Madison WI 53706; e-mail: huttenlocher@wisc.edu.
