

Key Points
RBC phagocytosis by neutrophils induces expression of MHC-II and costimulatory molecules and inhibits the respiratory burst.
Neutrophils can induce TT-specific T-cell activation and proliferation after RBC phagocytosis.

Abstract
Neutrophils are particularly well known for their antimicrobial function. Although historically they are regarded as strictly a phagocyte of the innate immune system, over time it has become clear that neutrophils are versatile cells with numerous functions including innate and adaptive immune regulation. We have previously described a role for human neutrophils in antibody-mediated red blood cell (RBC) clearance. Under homeostatic conditions, neutrophils do not take up RBCs. Yet, when RBCs are immunoglobulin G (IgG) opsonized, which can occur in alloimmunization or autoimmunization reactions, neutrophils can effectively phagocytose RBCs. In the present study, we show that human neutrophils acquire an antigen-presenting cell (APC) phenotype following RBC phagocytosis. Subsequent to RBC phagocytosis, neutrophils expressed major histocompatibility complex class II (MHC-II) and costimulatory molecules such as CD40 and CD80. Moreover, in classical APCs, the respiratory burst is known to regulate antigen presentation. We found that the respiratory burst in neutrophils is reduced after IgG-mediated RBC phagocytosis. Additionally, following RBC phagocytosis, neutrophils were demonstrated to elicit an antigen-specific T-cell response, using tetanus toxoid (TT) as an antigen to elicit an autologous TT-specific CD4+ T-cell response. Lastly, although the “don’t eat me” signal CD47 is known to have a powerful restrictive role in the activation of immunity against RBCs in dendritic cells, CD47 does not seem to have a significant effect on the antigen-presenting function of neutrophils in this context. Overall, these findings reveal that besides their classical antimicrobial role, neutrophils show plasticity in their phenotype.
Introduction
Neutrophils are innate immune cells that are the first responders in tissue injury and infection.1,2 They were conventionally regarded as terminally differentiated cells with an antimicrobial function. Over the years, it has become clear that the function of neutrophils extends well beyond the classical role of an innate immune cell.3 It has been established that neutrophils possess a broad assortment of cytokines and effector molecules.4,5 In addition, neutrophils have been shown to be involved in an extensive range of effector functions and can activate and regulate the innate and adaptive immune system.3
In a previous study, we have described a role for neutrophils in antibody-mediated red blood cell (RBC) clearance.6 Because the spleen is the major filter of the blood and the primary organ responsible for RBC clearance, we focused on RBC clearance in this organ. We found that whereas homeostatic RBC clearance is mainly a task for splenic macrophages, neutrophils can become the primary phagocyte in the clearance of immunoglobulin G (IgG)–opsonized RBCs (RBC-ops). These findings suggest a role for neutrophils in autoimmune or alloimmune reactions against RBCs after the formation of the primary antibody. In the current study, we explored the consequences of RBC phagocytosis on immune functions of the neutrophil.
Increasing evidence indicates that neutrophils can contribute to adaptive immunity by influencing antigen-specific responses. They can have an indirect effect on antigen presentation by activating dendritic cells (DCs)7 and they may even directly activate T cells by transporting and presenting antigens themselves.3,8-10 In this study, we have explored the potential of human neutrophils to act as antigen-presenting cells (APCs) following IgG-mediated RBC phagocytosis.
To present antigens to T cells, APCs need to express major histocompatibility complex class II (MHC-II). Additionally, costimulatory molecules are necessary for T-cell activation and proliferation. Therefore, we first examined the potential of neutrophils to express MHC-II and costimulatory molecules. Next, we have investigated the respiratory burst that results from RBC phagocytosis. In professional APCs, the extent of reactive oxygen species (ROS) production helps to regulate the level of antigen degradation and thereby the efficiency of antigen presentation.11-13 Ultimately, we have investigated the ability of neutrophils to induce a specific T-cell response.
Under homeostatic conditions, 2.5 × 1011 RBCs become senescent and get cleared from the circulation each day.14 Virtually all cells including RBCs express CD47 as a marker of “self.”15 CD47 acts as a molecular switch for erythrophagocytosis16 and, additionally, CD47–signal-regulatory protein α (SIRPα) interactions negatively control various immune effector functions.17 Yi et al have demonstrated that reduced expression of CD47 activates DCs and contributes to autoimmunity or alloimmunity against RBCs.18 We have previously found that CD47-SIRPα interactions act as an inhibiting signal in erythrophagocytosis by neutrophils. Whether the lack of CD47 on RBCs can also promote the induction of antigen-specific CD4+ T-cell responses when using neutrophils as APCs is as yet unclear.
In this study, we show for the first time that the phagocytosis of IgG–RBC-ops causes human neutrophils to acquire APC characteristics such as the expression of MHC-II and costimulatory molecules. Moreover, we demonstrate that the respiratory burst is greatly reduced in neutrophils that phagocytose RBC-ops compared with neutrophils taking up microbes. Additionally, using tetanus toxoid (TT) as an antigen, these neutrophils were proven to elicit an autologous TT-specific CD4+ T-cell response. This T-cell response is not affected by CD47 on the RBCs taken up by the neutrophils. Overall, our findings show that neutrophils are versatile cells with a plastic phenotype.
Methods
Isolation of RBCs
RBCs were isolated as previously described16 from healthy donor blood, after obtaining informed consent. Studies on human blood samples were approved by the Sanquin Research institutional medical ethical committee, in accordance with the 1964 Declaration of Helsinki.
Isolation of human neutrophils
Neutrophils (polymorphonuclear neutrophils) were isolated as previously described.19 All blood was obtained after informed consent and according to the 1964 Declaration of Helsinki.
Antibodies
Anti-glycophorin A (anti-GPA) (CD235a, mouse monoclonal IgG1; Sanquin Reagents) was used to opsonize RBCs. GPA is the most prevalent protein on the RBC membrane and is therefore a good target for opsonization. In our previous paper, this antibody was found to be highly potent in inducing phagocytosis by neutrophils.6
F(ab′)2 anti-CD47 (mouse monoclonal IgG1, clone B6H12, generously provided by E. Brown, University of California, San Francisco, San Francisco, CA) was used to block CD47 on RBCs. Antibodies used for fluorescence-activated cell sorter (FACS) analysis were: anti-CD3 Pacific Blue (Life Technologies), anti-CD4 allophycocyanin (Becton, Dickinson & Company), anti-CD8 phycoerythrin (PE; BioLegend), anti-CD19 allophycocyanin (BD Pharmingen), anti-CD14 PE-Cy7 (BD Pharmingen), anti-HLA-DR Pacific Blue (BioLegend), anti-HLA-DR fluorescein isothiocyanate (FITC; BioLegend), anti-CD40 brilliant violet 510 (BioLegend), anti-CD40 FITC (BioLegend), anti-CD80 FITC (BioLegend), anti-CD83 allophycocyanin (BD Pharmingen), anti-CD86 FITC (R&D Systems), anti-CD25 BV510 (BD Biosciences), anti-CD38 PE-Cy7 (BD Biosciences), isotype FITC (Pelicluster), isotype allophycocyanin (Sanquin), isotype PE (Pelicluster), isotype PE-Cy7 (BD Biosciences), and isotype brilliant violet 510 (BD Biosciences).
Phagocytosis assay
RBCs were stained with DiD (1,1′-dioctadecyl-3,3,3′, 3′-tetramethylindodicarbocyanine 4-chlorobenzenesulfonate salt) lipophilic membrane dye (Life Technologies). Subsequently, RBCs plus or minus anti-GPA (RBC and RBC-ops) and plus or minus F(ab′)2 anti-CD47 were incubated with neutrophils isolated from blood for 1 hour or overnight at 37°C (10:1 ratio, 5 × 106 RBCs plus or minus 0.5 × 106 neutrophils). Next, RBCs were lysed using an isotonic ammonium chloride buffer (isotonic NH4Cl-KHCO3-EDTA solution) for 10 minutes at 4°C followed by a second lysis step of 5 minutes at 4°C. Cells were washed and resuspended in stop buffer containing 0.5% (wt/vol) paraformaldehyde, 1% (wt/vol) bovine serum albumin, and 20 mM NaF in phosphate-buffered saline. Phagocytosis was measured using a Canto II flow cytometer (BD Biosciences) equipped with FACSDiva software (BD Biosciences). For measurement of HLA-DR, CD40, and CD80 expression, the same procedure was performed with unstained neutrophils. Neutrophils in the various conditions were cultured overnight with 10 ng/mL granulocyte colony-stimulating factor (G-CSF; PeproTech), and HLA-DR, CD40, and CD80 expression was measured with flow cytometry on fresh neutrophils and after overnight incubation. Represented mean fluorescence intensity (MFI) shows MFI of stained neutrophils minus MFI of neutrophils with isotype control.
Phagocytosis assay; magnetic selection
RBCs were incubated in 15 μg/mL biotin (Thermo Scientific) in PBS at 37°C for 30 minutes and then washed and incubated with streptavidin MicroBeads (Miltenyi Biotec) for 30 minutes at 4°C. Hereafter, RBCs were opsonized with anti-GPA (30 minutes, 4°C). The magnetic RBC-ops were washed and added to freshly isolated neutrophils (10:1 ratio, 45 minutes at 37°C in N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid [HEPES] medium). Next, RBCs were lysed using an isotonic ammonium chloride buffer for 5 minutes at 4°C followed by a second lysis step of 5 minutes at 4°C. After washing, the phagocytic cells were selected using an LS magnetic-activated cell sorting column (Miltenyi Biotec). The phagocytic and nonphagocytic fraction were incubated overnight (37°C, HEPES medium with 10 ng/mL G-CSF) and analyzed for MHC-II and costimulatory molecule expression using Canto II (BD Biosciences) flow cytometry equipped with FACSDiva software (BD Biosciences). Represented MFIs show MFI of stained neutrophils minus MFI of neutrophils with isotype control.
Quantitative PCR
The QIAamp RNA isolation kit (Qiagen) was used for all RNA isolation according to the manufacturer’s protocol. For the generation of complementary DNA, the SuperScript III First Strand Synthesis System for reverse transcription polymerase chain reaction (PCR; Invitrogen) was used according to the manufacturer’s protocol. Reverse transcription PCR was performed on the Light Cycler system (Roche), using the following amplification conditions: 2 minutes at 95°C, 5 seconds at 98°C, 10 minutes at 60°C followed by 15 minutes at 72°C. Primers for HLA-DR and β-glucuronidase (GUS) (control) were as followed: HLA-DR exon 1 forward: CTC AGG AAT CAT GGG CTA ACA AAG; HLA-DR exon 2-3 reverse: TAC CTC TGG AGG TAC ATT GGT GAT; GUS forward: GAA AAT ATG TGG TTG GAG AGC TCA TT; GUS reverse: CCG AGT GAA GAT CCC CTT TTT A. The PCR product was checked by Sanger sequencing (Abiprism 3700 genetic analyzer; Applied Biosystems) and with Blast control on specificity.
DC culture
DCs were obtained from the culture of CD14 microbead (Miltenyi Biotec) isolated monocytes. The monocytes were seeded at a density of 1 × 106/mL in a 6-well culture plate in Cellgro DC medium (Cellgenix) containing 800 U/mL interleukin 4 (Cellgenix) and 1000 U/mL granulocyte-macrophage colony-stimulating factor (GM-CSF; Cellgenix). After a 7-day incubation at 37°C, immature DCs were harvested and stimulated for 48 hours with lipopolysaccharide (LPS; 1 µg/mL) in medium with 1% fetal calf serum. FACS analysis for MHC-II and costimulatory molecule expression was performed using a Canto II (BD Biosciences) equipped with FACSDiva software (BD Biosciences). Represented MFIs show MFI of stained neutrophils minus MFI of neutrophils with isotype control.
NADPH oxidase activity
The respiratory burst was measured with the Amplex Red (10-acetyl-3,7-dihydroxyphenoxazine) Hydrogen Peroxidase Assay kit (Molecular Probes) as described previously.20 Neutrophils were incubated plus or minus RBCs or RBC-ops. To inhibit catalase activity of RBCs, the assay was performed in the presence of NaN3 (2 mM; Merck). To control for the ability to produce ROS, the different neutrophil incubations were additionally incubated with either zymosan (1 mg/mL; Sigma-Aldrich) or phorbol 12-myristate 13-acetate (PMA) (100 ng/mL; Sigma-Aldrich). Fluorescence was measured at 30-second intervals for 30 minutes with the Infinite 200 PRO (Tecan). Maximal slope of H2O2 release was assessed over a 2-minute interval to determine the reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activity. Intracellular H2O2 production was measured by dihydrorhodamine-1,2,3 (DHR)-labeled RBCs or zymosan (29 mM; Sigma Chemicals) as described in detail previously.21
Nitroblue tetrazolium assay
Neutrophils were cultured in HEPES medium19 with 1 mg/mL G-CSF overnight at 37°C plus or minus RBCs or RBCs-ops or for 5 minutes with PMA. After incubation, nitroblue tetrazolium (NBT; Sigma) in PBS (5 mg/mL) was added to the cell suspensions and incubated at 37°C for 30 minutes. Cytospins were prepared using ∼100 000 cells and fixed using methanol. KernEchtRood (Merck) was added to slides for 5 to 10 minutes, which were then rinsed and inspected under a microscope.
Western blot
Lysates of fresh neutrophils and overnight incubated neutrophils plus or minus RBCs or RBC-ops plus or minus F(ab′)2 anti-CD47 were made and subjected to 10% sodium dodecyl-sulfate–polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes (Schleicher & Schuell). Membranes were washed and blocked with 5% nonfat milk (Elk; Campina). As primary antibody, anti–HLA-DR (mouse IgG; BioLegend) or anti-CD40 (mouse IgG; Novis) were used. As loading control, the housekeeping protein glyceraldehyde-3-phosphate dehydrogenase (GADPH) (rabbit IgG; Merck Millipore) or actin (Sigma) were detected (2% milk protein/Tris-buffered saline Tween 20 overnight at 4°C). As secondary antibody, goat anti–mouse-IgG IRDye 800CW or goat anti–rabbit-IgG IRDye 680RD (LI-COR Biosciences) were used. Quantification of protein expression was performed on an Odyssey Infrared Imaging system (LI-COR Biosciences).
Autologous TT-specific T-cell assay
Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood using density gradient centrifugation over isotonic Percoll (Pharmacia) with a specific gravity of 1.076 g/mL (2070 rpm, 20 minutes, 21°C). Next, magnetic-activated cell sorting isolation was used to isolate T cells (Pan T-cell isolation kit human; Miltenyi Biotec). Purity of isolated T cells and neutrophils was established with cytospin and flow cytometry (supplemental Figures 2 and 3). T cells were incubated with neutrophils plus or minus RBCs or RBC-ops (T cells to neutrophils to RBCs, 5:1:10 ratio) and 10 µg/mL TT (Statens Serum Institute). As a positive control, autologous T-cell–depleted PBMCs (plus or minus 10 µg/mL TT) were used to stimulate the T cells (T cells to PBMCs, 5:1). As a negative control, T cells alone and/or T cells with RBCs (5:10 ratio, 10 µg/mL TT) were used. In addition, to establish an antigen-specific effect, all conditions were also evaluated without TT. On day 4, T cells were restimulated with neutrophils plus or minus RBCs or RBC-ops. Activation of T cells was measured on days 1, 4, and 8 by measuring the percentage of CD25, CD38, or HLA-DR+ CD4+ or CD8+ cells by flow cytometry (Canto II; BD Biosciences). To measure T-cell proliferation, T cells were carboxyfluorescein diacetate succinimidyl ester (CFSE)-labeled (Cell Trace; Invitrogen) at day 0. Proliferation of T cells was measured on day 8 by measuring the percentage of CFSE dim CD4+ or CD8+ cells by flow cytometry. (Canto II; BD Biosciences). The gating strategy is explained in supplemental Figure 4. HLA-DR was blocked using HLA-DR–blocking antibody (BioLegend).
Luminex
Twenty-five microliters of medium from each condition of the TT-specific T-cell assay was analyzed using a custom Bio-Plex Pro human cytokine 13-plex panel antibody kit according to the manufacturer’s protocol (Bio-Rad). As a readout system, we used a Bio-Plex protein array system (Bio-Rad) using both supplied high-calibration and low-calibration curves, according to the manufacturer’s instructions.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 7. For comparison of phagocytosis, protein expression, H2O2 production, and proliferation, the 1-way analysis of variance test was used followed by the Sidak post hoc test for correction of multiple comparison. For the comparison of the expression levels of the activation markers, a 2-way analysis of variance test was used followed by the Tukey test for correction of multiple comparison (****P < .0001; ***P < .001; **P < .01; *P < .05).
Results
RBC phagocytosis by neutrophils induces MHC-II and costimulatory molecule expression
We have previously shown that only RBC-ops, and not unopsonized RBCs, are phagocytosed by neutrophils.6 To investigate whether neutrophils can act as APCs following RBC phagocytosis, we first confirmed our previous findings using flow cytometry and cytospin analysis (Figure 1A-B; supplemental Figure 1). Professional APCs express MHC-II and costimulatory molecules in order to present antigens. Therefore, we measured MHC-II (HLA-DR) and costimulatory molecule (CD40 and CD80) expression on neutrophils following RBC phagocytosis. Freshly isolated neutrophils showed no to low expression of HLA-DR, CD40, and CD80. In contrast, these markers were upregulated on neutrophils 20 hours after RBC phagocytosis (Figure 1C-E) (these results were confirmed for HLA-DR and CD40 by western blot [see Figure 6D-E]). Previously, interferon γ (IFNγ), tumor necrosis factor α (TNFα), and GM-CSF were shown to induce HLA-DR expression on neutrophils.22,23 Here, we found that the levels of HLA-DR but not CD40 or CD80 upregulation on neutrophils were significantly greater after incubation with RBC-ops when compared with neutrophils stimulated with IFNγ, TNFα, or GM-CSF (Figure 1F-H).
![Figure 1. Neutrophils upregulate MHC-II and costimulatory molecules following RBC phagocytosis. (A-B) RBC phagocytosis by neutrophils was measured by incubating neutrophils ± DiD-labeled unopsonized RBCs (RBC) or anti-GPA RBC-ops (RBC-ops) for 45 minutes. Subsequently, RBCs were lysed and the percentage of RBC-phagocytosing neutrophils was measured by flow cytometry. (A) Approximately 45% of neutrophils phagocytose RBC-ops (mean ± standard error of the mean [SEM]; n = 5). (B) Phagocytosis of RBC-ops by neutrophils was confirmed by cytospin. These images where blindly chosen and are representative for 5 different experiments. May-Grünwald Giemsa stain; original magnification ×500. (C-E) Surface expression of HLA-DR (MHC-II), CD40, and CD80 on freshly isolated neutrophils (T = 0) and neutrophils cultured for 20 hours (T = 20) with or without RBCs or RBC-ops. (F-H) Surface expression of HLA-DR, CD40, and CD80 on freshly isolated neutrophils (T = 0) and neutrophils stimulated with RBC-ops, IFNγ, TNFα, or GM-CSF and cultured for 20 hours (T = 20). The data represent the MFI values of HLA-DR, CD40, and CD80 (mean ± SEM; n = 5-7). Asterisks above straight-line bars represent significant differences compared with T = 0 control; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; ***P < .001; **P < .01; *P < .05). IFN, interferon; ns, nonsignificant.](https://ash.silverchair-cdn.com/ash/content_public/journal/bloodadvances/3/11/10.1182_bloodadvances.2018028753/2/m_advances028753f1.png?Expires=1763494657&Signature=K8Bkj8Ul8KE21nkbJJessa7xFDm4tmz8r16phi8~zWfV8h4QvJvEE9D1OB2l5VNcxT1dragZzqJDFG~eVTcBdFclmn9rwIMrpzJ6ONzvGS-OZHfo7fFjfyeGv18g6CELQO~3TRmmBm7ZtW88o8Gs7-MvUZlMwZ01tEisKFcj2ARibBVao0hFYM4bSDG64okmFgIU5UB5ZbAKweRNRP-ssrlu0i5RN8s71bOnMbgrhQ9aFRxZmVV9i0xcjgTWhnw6W-Apu3WusaHgd04QU8Tv98wBt-9Vrl3siWJBw2k6gRcbomGo40h~leCs1sboplb8~nicGbi-SAVvFz5E0z9RDw__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Neutrophils upregulate MHC-II and costimulatory molecules following RBC phagocytosis. (A-B) RBC phagocytosis by neutrophils was measured by incubating neutrophils ± DiD-labeled unopsonized RBCs (RBC) or anti-GPA RBC-ops (RBC-ops) for 45 minutes. Subsequently, RBCs were lysed and the percentage of RBC-phagocytosing neutrophils was measured by flow cytometry. (A) Approximately 45% of neutrophils phagocytose RBC-ops (mean ± standard error of the mean [SEM]; n = 5). (B) Phagocytosis of RBC-ops by neutrophils was confirmed by cytospin. These images where blindly chosen and are representative for 5 different experiments. May-Grünwald Giemsa stain; original magnification ×500. (C-E) Surface expression of HLA-DR (MHC-II), CD40, and CD80 on freshly isolated neutrophils (T = 0) and neutrophils cultured for 20 hours (T = 20) with or without RBCs or RBC-ops. (F-H) Surface expression of HLA-DR, CD40, and CD80 on freshly isolated neutrophils (T = 0) and neutrophils stimulated with RBC-ops, IFNγ, TNFα, or GM-CSF and cultured for 20 hours (T = 20). The data represent the MFI values of HLA-DR, CD40, and CD80 (mean ± SEM; n = 5-7). Asterisks above straight-line bars represent significant differences compared with T = 0 control; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; ***P < .001; **P < .01; *P < .05). IFN, interferon; ns, nonsignificant.
Neutrophils upregulate MHC-II and costimulatory molecules following RBC phagocytosis. (A-B) RBC phagocytosis by neutrophils was measured by incubating neutrophils ± DiD-labeled unopsonized RBCs (RBC) or anti-GPA RBC-ops (RBC-ops) for 45 minutes. Subsequently, RBCs were lysed and the percentage of RBC-phagocytosing neutrophils was measured by flow cytometry. (A) Approximately 45% of neutrophils phagocytose RBC-ops (mean ± standard error of the mean [SEM]; n = 5). (B) Phagocytosis of RBC-ops by neutrophils was confirmed by cytospin. These images where blindly chosen and are representative for 5 different experiments. May-Grünwald Giemsa stain; original magnification ×500. (C-E) Surface expression of HLA-DR (MHC-II), CD40, and CD80 on freshly isolated neutrophils (T = 0) and neutrophils cultured for 20 hours (T = 20) with or without RBCs or RBC-ops. (F-H) Surface expression of HLA-DR, CD40, and CD80 on freshly isolated neutrophils (T = 0) and neutrophils stimulated with RBC-ops, IFNγ, TNFα, or GM-CSF and cultured for 20 hours (T = 20). The data represent the MFI values of HLA-DR, CD40, and CD80 (mean ± SEM; n = 5-7). Asterisks above straight-line bars represent significant differences compared with T = 0 control; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; ***P < .001; **P < .01; *P < .05). IFN, interferon; ns, nonsignificant.
Approximately 45% of the neutrophils incubated with RBC-ops phagocytozed RBCs (Figure 1A). To selectively analyze the subset of neutrophils that had phagocytozed an RBC, we developed a highly specific assay using magnetically labeled RBCs (Figure 2A). Using magnetic selection, we separated nonphagocytizing neutrophils from phagocytosing neutrophils (Figure 2B) and we measured MHC-II and costimulatory molecule expression on both subsets using flow cytometry. HLA-DR, CD40, and CD80 were significantly upregulated on phagocytosing neutrophils but remained negative on nonphagocytosing neutrophils compared with the control (Figure 2C-E). Moreover, due to the selectivity of this assay, the MFI for the markers is greatly increased compared with the standard flow cytometric assay used in Figure 1C-H. In addition, we determined CD86 and CD83 expression on the different sets of neutrophils (Figure 2F-G). These markers showed no or very low expression on freshly purified blood neutrophils, as well as on nonphagocytosing or phagocytosing neutrophils. Finally, we found that messenger RNA levels for HLA-DR were also significantly upregulated in phagocytosing neutrophils (Figure 2H). These results show that neutrophils selectively acquire characteristics of an APC following IgG-opsonized RBC phagocytosis.
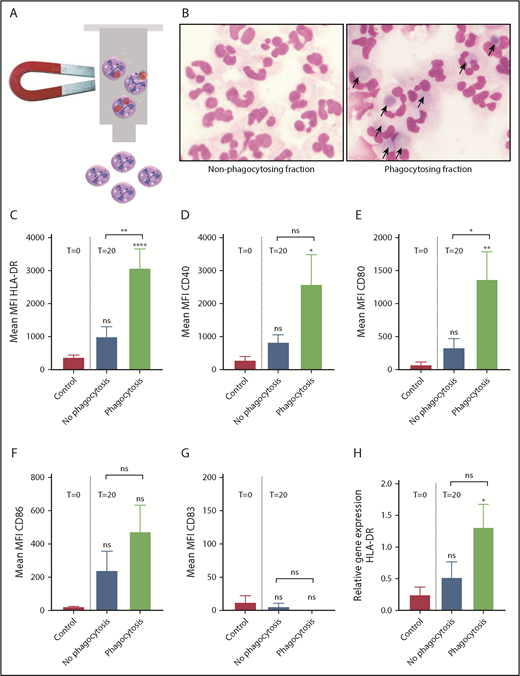
MHC-II and costimulatory molecules are specifically upregulated on neutrophils that have phagocytized RBCs. (A) Schematic overview showing magnetic selection of the neutrophils that have phagocytosed RBC-ops. RBCs were biotinylated and coupled to magnetic streptavidin beads. RBCs were opsonized and incubated with neutrophils for 1 hour. Next, RBCs were lysed and phagocytosing neutrophils were obtained by magnetic selection. (B) Cytospins of the nonphagocytosing and phagocytosing neutrophils after magnetic separation. These images were blindly chosen and are representative for 5 different experiments. May-Grünwald Giemsa stain; original magnification ×500. (C-G) Surface expression of HLA-DR, CD40, and CD80 on nonphagocytosing and phagocytosing neutrophils. Data represent the MFI values of HLA-DR, CD40, and CD80 (mean ± SEM; n = 4-10). (H) Relative levels of mRNA for HLA-DR in neutrophils ± RBC-ops measured by quantitative PCR. Data are normalized for GUS expression (mean ± SEM; n = 4). Asterisks above the straight-line bars represent significant differences compared with T = 0 control; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; **P < .01; *P < .05).
MHC-II and costimulatory molecules are specifically upregulated on neutrophils that have phagocytized RBCs. (A) Schematic overview showing magnetic selection of the neutrophils that have phagocytosed RBC-ops. RBCs were biotinylated and coupled to magnetic streptavidin beads. RBCs were opsonized and incubated with neutrophils for 1 hour. Next, RBCs were lysed and phagocytosing neutrophils were obtained by magnetic selection. (B) Cytospins of the nonphagocytosing and phagocytosing neutrophils after magnetic separation. These images were blindly chosen and are representative for 5 different experiments. May-Grünwald Giemsa stain; original magnification ×500. (C-G) Surface expression of HLA-DR, CD40, and CD80 on nonphagocytosing and phagocytosing neutrophils. Data represent the MFI values of HLA-DR, CD40, and CD80 (mean ± SEM; n = 4-10). (H) Relative levels of mRNA for HLA-DR in neutrophils ± RBC-ops measured by quantitative PCR. Data are normalized for GUS expression (mean ± SEM; n = 4). Asterisks above the straight-line bars represent significant differences compared with T = 0 control; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; **P < .01; *P < .05).
To get more insight into the potential of neutrophils to perform a role as APC, we compared MHC-II and costimulatory molecule expression of RBC-ops–stimulated neutrophils with immature and mature monocyte-derived DCs and monocytes (Figure 3). Mature DCs express considerably higher levels of HLA-DR (288 times higher), CD40 (17.9 times higher), and CD80 (92.4 times higher) than neutrophils incubated with RBC-ops. CD40 is significantly higher on immature DCs and a trend of higher expression on immature DCs can be seen for HLA-DR and CD80 (HLA-DR, 179 times higher; CD40, 2.6 times higher; and CD80, 5.8 times higher). Due to large variation in expression of HLA-DR and CD80 between donors, expression levels for these proteins on immature DCs are not significantly enhanced compared with neutrophils incubated with RBC-ops. Interestingly, HLA-DR, CD40, and CD80 expression on neutrophils incubated with RBC-ops is comparable to the expression of these molecules on nonactivated monocytes.
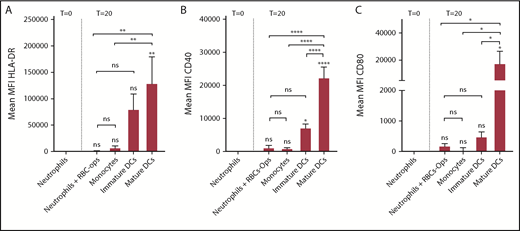
Mature DCs have far greater expression levels of MHC-II and costimulatory molecules. Surface expression of HLA-DR, CD40, and CD80 on freshly isolated neutrophils (T = 0), neutrophils plus RBC-ops cultured for 20 hours (T = 20), monocytes, immature or mature DCs. Data represent the MFI values of HLA-DR (A), CD40 (B), and CD80 (C) (mean ± SEM; n = 3-5). Asterisks above the straight-line bars represent significant differences compared with T = 0 neutrophils; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; **P < .01; *P < .05).
Mature DCs have far greater expression levels of MHC-II and costimulatory molecules. Surface expression of HLA-DR, CD40, and CD80 on freshly isolated neutrophils (T = 0), neutrophils plus RBC-ops cultured for 20 hours (T = 20), monocytes, immature or mature DCs. Data represent the MFI values of HLA-DR (A), CD40 (B), and CD80 (C) (mean ± SEM; n = 3-5). Asterisks above the straight-line bars represent significant differences compared with T = 0 neutrophils; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; **P < .01; *P < .05).
The respiratory burst is almost absent in neutrophils during RBC phagocytosis
As a rule, phagocytosis by neutrophils leads to the induction of a respiratory burst. To examine the consequence of RBC phagocytosis by neutrophils, we measured the respiratory burst following erythrophagocytosis. Surprisingly, the respiratory burst following RBC-ops uptake, quantified by measuring extracellular H2O2 production, was not enhanced compared with control neutrophils or neutrophils incubated with unopsonized RBCs (Figure 4A). Phagocytosis of yeast-derived zymosan particles or the addition of PMA, both potent inducers of the respiratory burst, showed that neutrophils in all 3 conditions were capable of producing ROS (Figure 4A). Similar results were obtained with the DHR assay, a FACS-based method that detects intracellular H2O2 production (Figure 4B). No burst was measurable for neutrophils incubated with RBC-ops when tested 180 minutes after the start of the phagocytosis assay. PMA was added to confirm that neutrophils in all 4 conditions were able to produce a respiratory burst.
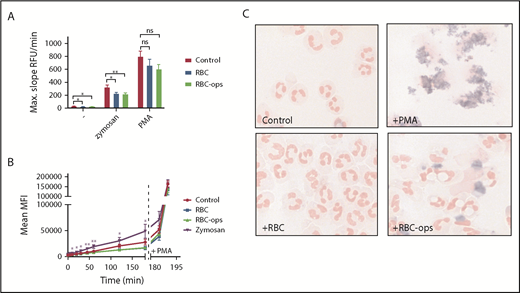
Phagocytosis of RBCs by neutrophils induces a strongly diminished respiratory burst. (A) Release of H2O2 was measured by Amplex Red assay for neutrophils incubated ± RBC or RBC-ops. No burst was measured in the 3 conditions (mean ± SEM; n = 10-12). To control for the ability to produce a respiratory burst, neutrophils from all 3 conditions were stimulated with zymosan or PMA (mean ± SEM; n = 10-12). (B) Release of intracellular H2O2 production was measured using a DHR assay with neutrophils incubated ± RBC or RBC-ops. Neutrophils incubated with zymosan served as a positive control. Addition of PMA at 180 minutes showed neutrophils in all conditions were capable of producing ROS (mean ± SEM; n = 4-13). Asterisks above data points represent significant differences for zymosan compared with control (**P < .01; *P < .05). (C) Respiratory burst was visualized using NBT, which produces a blue precipitate in presence of intracellular ROS. Neutrophils were incubated plus RBC or RBC-ops. Neutrophils plus PMA served as a positive control and neutrophils alone served as a negative control. Images are representative for 3 individual experiments. May-Grünwald Giemsa stain; original magnification ×500. RFU, relative fluorescence unit.
Phagocytosis of RBCs by neutrophils induces a strongly diminished respiratory burst. (A) Release of H2O2 was measured by Amplex Red assay for neutrophils incubated ± RBC or RBC-ops. No burst was measured in the 3 conditions (mean ± SEM; n = 10-12). To control for the ability to produce a respiratory burst, neutrophils from all 3 conditions were stimulated with zymosan or PMA (mean ± SEM; n = 10-12). (B) Release of intracellular H2O2 production was measured using a DHR assay with neutrophils incubated ± RBC or RBC-ops. Neutrophils incubated with zymosan served as a positive control. Addition of PMA at 180 minutes showed neutrophils in all conditions were capable of producing ROS (mean ± SEM; n = 4-13). Asterisks above data points represent significant differences for zymosan compared with control (**P < .01; *P < .05). (C) Respiratory burst was visualized using NBT, which produces a blue precipitate in presence of intracellular ROS. Neutrophils were incubated plus RBC or RBC-ops. Neutrophils plus PMA served as a positive control and neutrophils alone served as a negative control. Images are representative for 3 individual experiments. May-Grünwald Giemsa stain; original magnification ×500. RFU, relative fluorescence unit.
Because it is improbable that the burst is completely absent, we finally used an NBT assay to determine whether we could detect a respiratory burst following RBC phagocytosis. Superoxide reduces NBT, which results in a dark blue precipitate. The respiratory burst can be visualized hereby but not properly quantified. And indeed, neutrophils incubated with RBC-ops showed a slight but consistent blue precipitate in the NBT assay whereas control neutrophils and neutrophils incubated with unopsonized RBCs did not (Figure 4C). Collectively, these findings indicate that the respiratory burst following RBC phagocytosis is greatly reduced in neutrophils but not absent.
Neutrophils can activate T cells and induce antigen-specific proliferation following RBC phagocytosis
We found that neutrophils acquire APC characteristics following RBC phagocytosis. We next wanted to investigate whether these neutrophils are also capable of stimulating T cells in an antigen-specific manner by studying the upregulation of the activation markers CD25, CD38, and HLA-DR on T cells over time and by measuring antigen-specific T-cell proliferation using an autologous TT-specific T-cell assay (Figure 5A). Furthermore, we investigate the capacity of neutrophils to process and present antigens because TT requires intracellular processing for presentation. We incubated fresh neutrophils ± RBCs or RBC-ops ± TT with (CFSE-labeled) T cells. T cells alone and/or T cells with RBCs were taken along as a negative control. T-cell–depleted PBMCs were added as an APC source to the T cells and served as a positive control. To establish that measured T-cell proliferation is TT specific, all conditions were also evaluated without TT. Because neutrophils are short-living cells, the T cells cocultured with neutrophils were restimulated with fresh neutrophils on day 4. On days 1, 4, and 8, the expression levels of CD25, CD38, and HLA-DR were measured with flow cytometry (Figure 5B-C; supplemental Figure 5). Our results showed that CD25 is upregulated on CD4+ T cells on day 8 after coincubation of T cells with neutrophils and RBC-ops. The positive control (T cells incubated with PBMCs) already shows CD25 upregulation on CD4+ T cells on day 4. CD38 and HLA-DR are upregulated on CD4+ at day 8 in both the positive control, as in the condition where T cells are incubated with neutrophils and RBC-ops. On CD8+ T cells, CD25 is not significantly upregulated by neutrophils plus RBC-ops and only slightly upregulated by PBMCs on day 8 (supplemental Figure 5A). CD38 and HLA-DR are upregulated on day 8 by neutrophils plus RBC-ops and by PBMCs but expression is lower compared with CD4+ (supplemental Figure 5B-C). These results show that neutrophils are capable of specifically activating T cells following RBC phagocytosis.
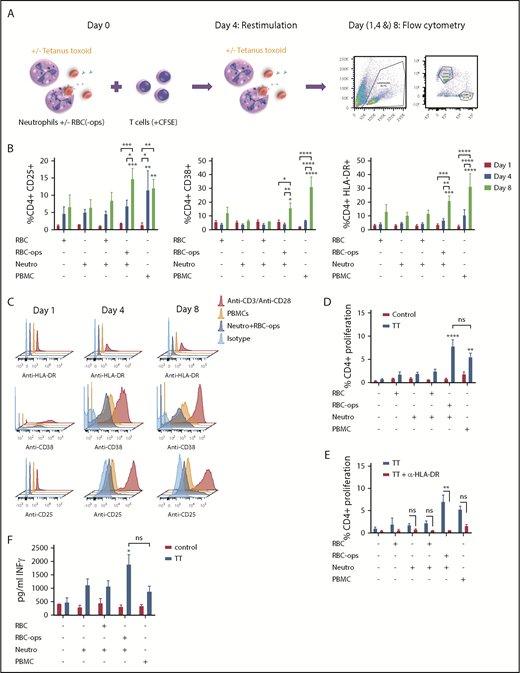
Neutrophils can activate T cells following RBC phagocytosis. (A) Schematic overview of the autologous TT-specific T-cell assay. On day 0, (CFSE-labeled) T cells were incubated with neutrophils ± RBCs or RBC-ops TT. On day 4, T cells were restimulated with neutrophils ± RBCs or RBC-ops and ± TT. Flow cytometry was used as readout on days (1, 4, and) 8. (B) T-cell activation was measured by flow cytometry by determining the percentage of CD25+, CD38, or HLA-DR+ CD4+ T cells after 1, 4, and 8 days. Results are shown for T cells incubated with neutrophils (Neutro) ± RBCs or RBC-ops. All conditions are plus TT. T cells plus RBCs served as a negative control and T cells plus PBMCs as positive control (mean ± SEM; n = 3). (C) Histograms showing CD25, CD38, and HLA-DR staining on CD4+ T cells on days 1, 4, and 8 of a representative donor. As a positive control for CD25, CD38 and HLA-DR staining T cells stimulated with anti-CD3 and anti-CD28 were used. An isotype control was used as negative control. (D) CD4+ T-cell proliferation was measured by flow cytometry by determining the percentage of CFSE low T cells after 8 days. Percentage proliferation is shown for T cells incubated with neutrophils ± RBCs or RBC-ops and ± TT. T cells ± RBCs served as a negative control and T cells plus PBMCs as positive control (mean ± SEM; n = 8). (E) The experiment was repeated with the addition of an MHC-II blocking antibody to show that neutrophil-induced T-cell proliferation is MHC-II restricted (mean ± SEM; n = 6). (F) Graph showing IFN-γ levels in the medium at day 8 (n = 6). Asterisks above the straight-line bars represent significant differences compared with T cells plus RBCs day 1; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; ***P < .001; **P < .01; *P < .05). Nonsignificant results have not been indicated.
Neutrophils can activate T cells following RBC phagocytosis. (A) Schematic overview of the autologous TT-specific T-cell assay. On day 0, (CFSE-labeled) T cells were incubated with neutrophils ± RBCs or RBC-ops TT. On day 4, T cells were restimulated with neutrophils ± RBCs or RBC-ops and ± TT. Flow cytometry was used as readout on days (1, 4, and) 8. (B) T-cell activation was measured by flow cytometry by determining the percentage of CD25+, CD38, or HLA-DR+ CD4+ T cells after 1, 4, and 8 days. Results are shown for T cells incubated with neutrophils (Neutro) ± RBCs or RBC-ops. All conditions are plus TT. T cells plus RBCs served as a negative control and T cells plus PBMCs as positive control (mean ± SEM; n = 3). (C) Histograms showing CD25, CD38, and HLA-DR staining on CD4+ T cells on days 1, 4, and 8 of a representative donor. As a positive control for CD25, CD38 and HLA-DR staining T cells stimulated with anti-CD3 and anti-CD28 were used. An isotype control was used as negative control. (D) CD4+ T-cell proliferation was measured by flow cytometry by determining the percentage of CFSE low T cells after 8 days. Percentage proliferation is shown for T cells incubated with neutrophils ± RBCs or RBC-ops and ± TT. T cells ± RBCs served as a negative control and T cells plus PBMCs as positive control (mean ± SEM; n = 8). (E) The experiment was repeated with the addition of an MHC-II blocking antibody to show that neutrophil-induced T-cell proliferation is MHC-II restricted (mean ± SEM; n = 6). (F) Graph showing IFN-γ levels in the medium at day 8 (n = 6). Asterisks above the straight-line bars represent significant differences compared with T cells plus RBCs day 1; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (****P < .0001; ***P < .001; **P < .01; *P < .05). Nonsignificant results have not been indicated.
Using the same assay, we determined the capacity of neutrophils to induce TT-specific T-cell proliferation (supplemental Figure 4). We found that neutrophils incubated with RBC-ops are capable of inducing CD4+ T-cell proliferation with a mean proliferation of 7.7%, comparable to proliferation induced by PBMCs (Figure 5D). Both neutrophils plus RBC-ops and PBMCs induce minimal CD8+ T-cell proliferation (supplemental Figure 5G). CD4+ T-cell proliferation induced by neutrophils is MHC-II restricted as demonstrated by using a blocking antibody for HLA-DR (Figure 5E). Lastly, after 8 days, IFN-γ secretion is significantly enhanced when T cells are incubated with neutrophils plus RBC-ops and TT compared with T cells incubated with RBCs (Figure 5F).
CD47 blocking on RBCs does not enhance the induction of TT-specific CD4+ T-cell proliferation
CD47 is a molecule that helps the immune system to discriminate self from nonself. This “don’t eat me” signal has a powerful restrictive role in the activation of DCs.18 To investigate whether this is also the case for neutrophils, we blocked CD47 on RBCs and RBCs-ops using a F(ab′)2 fragment against CD47 prior to incubation with neutrophils. As we have previously found, CD47 blocking augments phagocytosis of RBC-ops6 (Figure 6A). However, the increase in phagocytosis does not seem to affect antigen presentation by neutrophils. CD4+ T-cell proliferation was not significantly enhanced when neutrophils had taken up RBCs when CD47 was blocked although a trend is seen (Figure 6B). Additionally, CD47 blocking on RBC-ops does not seem to alter expression levels of HLA-DR, CD40, and CD80 on neutrophils after phagocytosis of these RBCs (Figure 6C). Western blot does show a slight increase in HLA-DR and CD40 expression, which could be explained by the fact that here we also measure intracellular protein. Overall, CD47 blocking on RBCs increases phagocytosis by neutrophils but does not seem to have a significant effect on the subsequent antigen-presenting function of neutrophils.
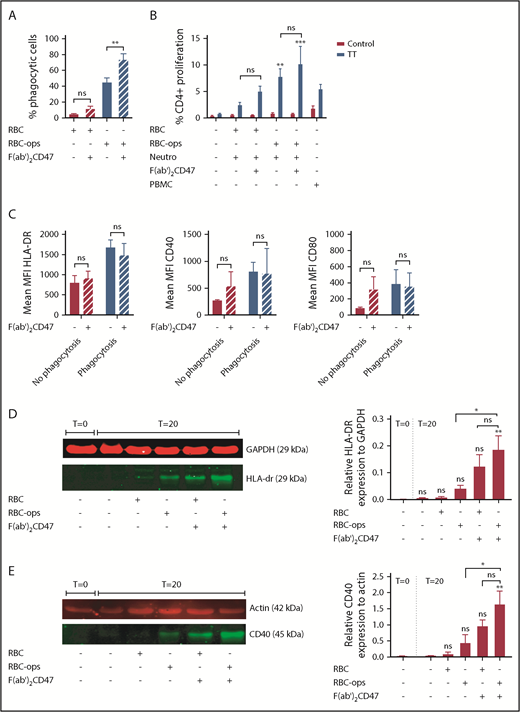
CD47 blocking on RBCs enhances the induction of TT-specific CD4+T-cell proliferation. (A) RBC phagocytosis by neutrophils was measured by incubating neutrophils ± DiD-labeled plus RBCs or RBC-ops, ± F(ab′)2 CD47 for 45 minutes. Subsequently, RBC were lysed and the percentage of RBC-phagocytosing neutrophils was measured by flow cytometry. Comparing the RBC-ops condition with the RBC-ops- F(ab′)2 CD47 condition, RBC phagocytosis increases from 45% to 73% (mean ± SEM; n = 5). (B) The percentage of proliferation measured for T cells incubated with neutrophils plus RBCs or RBC-ops, ± F(ab′)2 CD47 and ± TT. T cells served as a negative control and T cells plus PBMCs as positive control (mean ± SEM; n = 8). (C) Surface expression of HLA-DR, CD40, and CD80 on nonphagocytosing and phagocytosing neutrophils (mean ± SEM; n = 4). (D) Western blot analysis for HLA-DR expression performed on neutrophils ± RBCs or RBC-ops and ± F(ab′)2 CD47. Fluorescence intensity of the bands was quantified using Odyssey Imaging system and normalized for GAPDH expression. (E) Western blot analysis for CD40 expression performed on neutrophils ± RBCs or RBC-ops and ± F(ab′)2 CD47. Fluorescence intensity of the bands was quantified using Odyssey Imaging system and normalized for actin expression. (D-E) May-Grünwald Giemsa stain; original magnification ×500. Asterisks above the straight-line bars represent significant differences compared with T = 0 control neutrophils; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (**P < .01; *P < .05).
CD47 blocking on RBCs enhances the induction of TT-specific CD4+T-cell proliferation. (A) RBC phagocytosis by neutrophils was measured by incubating neutrophils ± DiD-labeled plus RBCs or RBC-ops, ± F(ab′)2 CD47 for 45 minutes. Subsequently, RBC were lysed and the percentage of RBC-phagocytosing neutrophils was measured by flow cytometry. Comparing the RBC-ops condition with the RBC-ops- F(ab′)2 CD47 condition, RBC phagocytosis increases from 45% to 73% (mean ± SEM; n = 5). (B) The percentage of proliferation measured for T cells incubated with neutrophils plus RBCs or RBC-ops, ± F(ab′)2 CD47 and ± TT. T cells served as a negative control and T cells plus PBMCs as positive control (mean ± SEM; n = 8). (C) Surface expression of HLA-DR, CD40, and CD80 on nonphagocytosing and phagocytosing neutrophils (mean ± SEM; n = 4). (D) Western blot analysis for HLA-DR expression performed on neutrophils ± RBCs or RBC-ops and ± F(ab′)2 CD47. Fluorescence intensity of the bands was quantified using Odyssey Imaging system and normalized for GAPDH expression. (E) Western blot analysis for CD40 expression performed on neutrophils ± RBCs or RBC-ops and ± F(ab′)2 CD47. Fluorescence intensity of the bands was quantified using Odyssey Imaging system and normalized for actin expression. (D-E) May-Grünwald Giemsa stain; original magnification ×500. Asterisks above the straight-line bars represent significant differences compared with T = 0 control neutrophils; asterisks above the inverted U–shaped spanner bars represent significant differences between the indicated bars (**P < .01; *P < .05).
Discussion
In the current study, we investigated whether neutrophils can contribute to adaptive immunity after acquiring antigen-presentation capacity following RBC phagocytosis. We established that neutrophils express MHC-II and costimulatory molecules after RBC uptake and found a reduced respiratory burst in neutrophils that phagocytose RBCs. These neutrophils can induce autologous TT-specific CD4+ T-cell activation and proliferation to a similar extent as PBMCs. Additionally, we found that CD47-SIRPα interaction may have some restricting activity for HLA-DR upregulation on neutrophils. However, this negative feedback signal does not seem to have a significant effect on T-cell activation and proliferation.
It has previously been shown that human neutrophils acquire APC characteristics under the influence of stimuli such as GM-CSF, IFNγ, or interleukin 4.22-24 A few studies have also shown that stimulated neutrophils can induce antigen-specific T-cell proliferation, for instance by presenting bacterial or viral antigens.24-27 In the current study, we show for the first time that the phagocytosis of IgG-opsonized endogenous material drives neutrophils toward a functional APC phenotype.
Evidence for an in vivo role of neutrophils as APCs has also been described in literature. For instance, clinical studies have demonstrated that IFN-γ and GM-CSF stimulate upregulation of HLA-DR expression.28,29 Moreover, in patients suffering from autoimmune diseases such as Wegener granulomatosis and rheumatoid arthritis, neutrophils show elevated levels of MHC-II, CD80, and CD86 compared with healthy controls.30 In mice, neutrophils have been shown to act as APCs in a chronic colitis model.31 Furthermore, at inflammatory sites, murine neutrophils can differentiate into APCs that rapidly clear bacteria and present bacterial antigens to T cells.32 Recently, Vono et al used rhesus macaques to demonstrate that antigen-positive neutrophils in draining lymph nodes are capable of antigen presentation and induction of CD4+ T-cell proliferation after vaccination.27
We found that neutrophils induce T-cell proliferation to a comparable extent as PBMCs. In the PBMC mixture, monocytes and B cells fulfill the role of APC. Vono et al have compared human neutrophils with classical APCs (CD16+ monocytes, CD14+ monocytes, plasmacytoid DCs, and myeloid DCs) in respect to inducing specific T-cell proliferation and found a lower but consistent ability for neutrophils to induce proliferation using a cytomegalovirus pp65–specific or influenza hemagglutinin-specific assay.27 Other studies have compared neutrophils with monocytes in a TT-specific assay. The outcome of these studies varied with some studies showing a lower capacity to induce proliferation25 whereas others showed neutrophils to be superior to monocytes.24 Moreover, there is opposing evidence suggesting that neutrophils can have an inhibiting effect on T-cell activation and proliferation. A suppressive role of neutrophils has been well defined in cancer. Here, neutrophils, also known as granulocytic myeloid–derived suppressor cells, are able to suppress antitumor immune responses by inhibiting cytotoxic T-cell function.33,34 Besides neutrophils, immature and mature monocyte myeloid–derived suppressor cells can also act as suppressors of antitumor immune responses.33
These seemingly contradicting effects of neutrophils on adaptive immune responses suggest a regulatory role. Increasing evidence is found supporting the presence of multiple neutrophil phenotypes.35-38 These different phenotypes could arise through specific differentiation programs in the bone marrow or they may be induced by extracellular signals such as cytokines and chemokines.34 Depending on the environmental conditions, neutrophils can exhibit different regulatory functions. For instance, they produce cytokines and chemokines vital for the initiation of an inflammatory response,2 yet, they can also secrete anti-inflammatory factors that resolve inflammation.3 Neutrophils can induce DC maturation and activation,7,39,40 however, an inhibitory effect on DCs has also been described.41-45 The wide range of functions demonstrates not only an effector role but also a regulatory role for neutrophils.
Neutrophils and professional APCs such as macrophages, monocytes, and DCs share a common origin. Although developing from a common precursor, the various myeloid cells differentiate and acquire distinct and often complementary characteristics such as different tissue localization, different capacities to fight pathogens, and different levels of cytokine production. The combination of complementary and overlapping features of myeloid cells helps to fine-tune effector functions and results in sophisticated orchestration of the innate and adaptive immune system.46 Although neutrophils will most likely be unable to replace or take over APC functions of professional APCs, these abundant cells may still be crucial as APC in specific situations.
We have previously shown that CD47-SIRPα interaction restricts RBC phagocytosis by neutrophils.6 In the current study, we have found that this negative feedback system does not have a significant effect on the induction of neutrophil-activated antigen-specific CD4+ T-cell responses. Although not significant, we do find a slight enhancement in T-cell proliferation when blocking CD47. Flow cytometry showed that the expression of HLA-DR, CD40, and CD80 does not increase, although an increase of CD40 and CD80 when CD47 is blocked is found on western blot, suggesting an intracellular pool of the proteins.
In this study, we determined the role of neutrophils in IgG-mediated RBC clearance, which may give us insight into the role of neutrophils in RBC alloimmunization. We have investigated the situation in which the alloantibody is already present. Therefore, our work does not clarify the role of neutrophils in the formation of a primary alloantibody. However, we show that once an anti-RBC antibody is formed, neutrophils participate in RBC clearance and subsequently acquire APC characteristics. Although DCs are most likely more potent APCs and crucial for the initial phase of alloimmunization, neutrophils could be involved in sustaining the alloimmune reaction. Moreover, our findings support a role for neutrophils in the formation of secondary antibodies. Although neutrophils express relatively low amounts of MHC-II and costimulatory molecules their abundance can compensate for this and increase their impact as an APC.
Nonetheless, in vivo studies should be performed in future research to establish the exact contribution of neutrophils in alloimmunization. Moreover, it should be noted that we have used a mouse IgG1 antibody for RBC opsonization. Antibody specificity, subclass, and/or glycosylation may affect the ability of the phagocytosing neutrophils to function as an APC.
In conclusion, this paper primarily highlights the plasticity of neutrophils that were previously thought of as terminally differentiated cells with an antimicrobial function. Additionally, our results suggest an unexpected role for neutrophils in RBC alloimmunization. Overall, neutrophils appear to have a more prominent role in adaptive immune responses such as alloimmunization than previously anticipated.
The full-text version of this article contains a data supplement.
Acknowledgments
The authors thank all blood donors for their single and/or serial donations. Furthermore, they thank Krystalyn Hudson from the University of Washington School of Medicine for the fruitful discussions on this topic and insightful comments on their work.
S.M.M. was supported by a grant from the Dutch Ministry of Health awarded to T.K.v.d.B.
Authorship
Contribution: S.M.M. performed the experiments, analyzed the data, and wrote the manuscript; G.B. and S.v.W. performed and analyzed experiments; J.G. performed and supervised quantiative PCR experiments; A.S. and A.t.B. provided the protocol for DC culturing and helped design and interpret T-cell assays; R.v.B. and S.M.M. designed the study; R.v.B. and T.K.v.d.B. supervised the study, edited the manuscript, and served as principal investigators; and B.M.B., M.H.J., and T.W.K. revised and approved the final manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Robin van Bruggen, Department of Blood Cell Research, Sanquin Research and Laboratory Services, Plesmanlaan 125, 1066CX Amsterdam, The Netherlands; e-mail: r.vanbruggen@sanquin.nl.

