

Abstract
Background
Mutations in MYD88 are highly recurring in Waldenstrom's Macroglobulinemia (WM) patients and are important for establishing the diagnosis of WM. The most common mutation in MYD88 is c.978T>C resulting a proline substitution for leucine at amino acid position 265 (p.Leu265Pro). Both allele specific PCR (AS-PCR) and clinical diagnostic next generation sequencing (NGS) panels are used to detect mutated MYD88, though they differ in sensitivity and scope. In this study we screened 734 patients with WM by AS-PCR for MYD88 c.978T>C MYD88 followed by Sanger sequencing to clarify negative results for non-MYD88 p.Leu265Pro mutations and compared the findings to clinical NGS panel data from the same biopsy when available. We also investigated MYD88 isoform dysregulation and isoform specific effects of the observed mutations that may impact mutated MYD88 regulation which has not been previously studied in WM.
Methods
DNA from CD19-selected bone marrow mononuclear cells (BMMC) of 734 WM patients were used for the MYD88 c.978T>C AS-PCR assay previously described by us (Xu et al, Blood 2013). For patients wild-type for MYD88 c.978T>C by AS-PCR, Sanger sequencing of the open reading frame of MYD88 was performed for both DNA and RNA simultaneously isolated from CD19-selected BMMC. DNA was also used to validate the presence of c.978T>C by Sanger. Findings were compared to 222/734 (30.2%) patients who also underwent illumina miSeq based targeted next generation sequencing on a clinical diagnostic platform using unselected BMMC. NGS isoform specific expression estimates were calculated using Salmon for 77 WM patients and 34 healthy donors (Hunter et al, Blood 2016).
Results
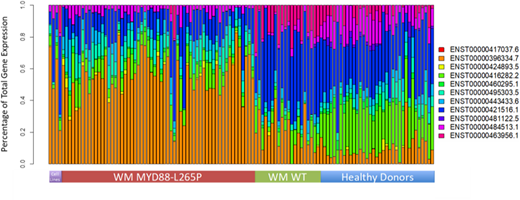
688/734 (93.7%) WM patients tested positive for the c.978T>C mutation. To confirm these results, Sanger sequencing at the DNA level covering the c.978T>C mutation was performed in 361/688 (52.5%) patients confirming the presence of the mutation in all cases. These Sanger studies revealed that one patient had two somatic mutations in addition to c.978T>C. Of the 46/734 (6.3%) that were wild-type by AS-PCR, 18 had cDNA available to screen for alternative MYD88 mutations. Of these, 13/18 (72.2%) were confirmed to be truly wild type for MYD88, and 5/18 (27.8%) harbored alternative MYD88 mutations making up 0.7% of the study population. Taken together 693/734 (94.4%) of patients were found to harbor somatic MYD88 mutations. Of the 222 patients form whom matching NGS panel data was available, the finding between the NGS and AS-PCR studies were largely concordant. The only discrepancies observed were 69 (31.1%) cases where targeted NGS gave false negative results for c.978T>C but was detected by AS-PCR. Of the four patients with alternative MYD88 mutations, one patient had a dinucleotide substitution that also resulted in p.Leu265Pro but tested as wild-type by AS-PCR, two patients each had one previously documented mutation (either pVal217Phe or p.Ser243Asn) and one patient had a mutation that was synonymous at the protein level (p.Phe277Phe). The patient with the two novel mutations in addition to c.978T>C had a mutation in the polypyrimidine track leading to the final exon and one resulting in p.Gly259Gly in the primary transcript but presents as a highly disruptive p.Val199Glu in the shorter regulatory isoforms. This is similar to c.978T>C which presents as p.L265P in the primary transcripts but acts as a stop loss in the shorter isoforms. We therefore looked for evidence of isoform level dysregulation in MYD88 using RNASeq and found highly significant and distinctive MYD88 isoform signatures for MYD88 mutant, MYD88 wild-type and healthy donor samples (Figure 1).
Conclusions
Using CD19-selected BMMC, MYD88 c.978T>C (p.Leu265Pro) was found in 93.7% of 734 patients, while non-c.978T>C mutations were present in <1% of WM population. Discordance with targeted NGS is common, with a false negative rate of 31%. Distinct isoform usage patterns and isoform specific mutational effects among MYD88 mutated patients was observed, highly suggestive of a functional role for MYD88 splicing in WM pathogenesis.
Castillo:Millennium: Research Funding; Janssen: Consultancy, Research Funding; Pharmacyclics: Consultancy, Research Funding; Abbvie: Consultancy, Research Funding; Genentech: Consultancy; Beigene: Consultancy, Research Funding. Treon:Johnson & Johnson: Consultancy; Janssen: Consultancy, Other: Travel, Accommodations, Expenses; BMS: Research Funding; Pharmacyclics: Consultancy, Other: Travel, Accommodations, Expenses, Research Funding. Hunter:Pharmacyclics: Consultancy.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal