Key Points
Newly acquired NANGS are significantly reduced in advanced-stage but not in early-stage t(14;18)-negative FL.
IGHV4-34 usage in combination with newly acquired NANGS in FR3 are significantly associated with t(14;18)-negative FL.

Abstract
We previously reported that t(14;18)-negative follicular lymphomas (FL) show a clear reduction of newly acquired N-glycosylation sites (NANGS) in immunoglobulin genes. We therefore aimed to investigate in-depth the occurrence of NANGS in a larger cohort of t(14;18)-positive and t(14;18)-negative FL, including early (I/II) and advanced (III/IV) stage treatment-naive and relapsed tumors. The clonotype was determined by using a next-generation sequencing approach in a series of 68 FL with fresh frozen material [36 t(14;18) positive and 32 t(14;18) negative]. The frequency of NANGS differed considerably between t(14;18)-positive and t(14;18)-negative FL stage III/IV, but no difference was observed among t(14;18)-positive and t(14;18)-negative FL stage I/II. The introduction of NANGS in all t(14;18)-negative clinical subgroups occurred significantly more often in the FR3 region. Moreover, t(14;18)-negative treatment-naive FL, specifically those with NANGS, showed a strong bias for IGHV4-34 usage compared with t(14;18)-positive treatment-naive cases with NANGS; IGHV4-34 usage was never recorded in relapsed FL. In conclusion, subgroups of t(14;18)-negative FL might use different mechanisms of B-cell receptor stimulation compared with the lectin-mediated binding described in t(14;18)-positive FL, including responsiveness to autoantigens as indicated by biased IGHV4-34 usage and strong NANGS enrichment in FR3.
Introduction
Follicular lymphoma (FL) is an indolent but incurable germinal center (GC) B-cell lymphoma.1 The median overall survival of patients with this disease is >15 years; however, ∼20% of patients progress or relapse within the first 2 years after initiation of treatment.2 The translocation t(14;18)(q32;q21), leading to a deregulated expression of the antiapoptotic protein BCL2, is the genetic hallmark of FL.1 It is found in ∼90% of advanced-stage (stage III/IV) and ∼50% of early-stage (stage I/II) FL.3 BCL2 expression, detectable in the majority of both t(14;18)-positive and t(14;18)-negative FL, either prolongs the stay of B cells in the GC or, rather, allows an iterative re-entry of t(14;18)-positive memory B cells to the GC,2,4 where they are constantly exposed to somatic hypermutation (SHM). This situation leads to an accumulation of secondary genetic hits and generates lymphoma precursor cells that eventually give rise to FL.2 Despite the disruption of one immunoglobulin (Ig) allele caused by the t(14;18), the B-cell receptor (BCR) remains functional in most FL, thus suggesting a specific role in this malignancy.5 SHM is also responsible for the introduction of newly acquired N-glycosylation sites (NANGS) (defined as an acquired Asn-X-Ser/Thr amino acid [AA] motif) in the Ig protein sequence in 80% to 100% of FL,5,6 which were found to be early and stable events in FL pathogenesis, despite ongoing SHM.7 In contrast to the more common N-glycosylations that occur at the Ig constant regions and include complex sugars, NANGS in FL are generated at the immunoglobulin variable heavy chain (IGHV) region, predominantly in the complement-determining region 2 (CDR2), and are of the high-mannose type.8 Notably, these NANGS are less common in other GC-derived lymphoma and rare in post–GC-derived lymphoma.5 In addition to these NANGS, there are naturally occurring potential NANGS in the CDR2 of the IGHV4-34 gene.8 Compared with NANGS, however, these natural sites lack glycosylation and the capability of binding to lectins expressed by M2 macrophages.9
Another feature of FL is its dependence on the microenvironment, which is composed of macrophages, follicular dendritic cells, and several T-cell subsets.10-15 Interestingly, higher numbers of macrophages and dendritic cells expressing lectins, capable of stimulating the BCR,16 were described as being associated with inferior prognosis.10,12,15,17
Of note, most published studies on the subject included random FL cohorts and were mainly composed of t(14;18)-positive stage III/IV FL, which represent the majority of cases in routine clinical practice. Therefore, little is known about N-glycosylation in t(14;18)-negative and stage I/II FL so far. In a recent study we found that, compared with published historical FL cohorts, NANGS were present in only one of six t(14;18)-negative FL (17% vs 80% to 100%).18 To better characterize t(14;18)-negative stage III/IV FL and stage I/II FL, which are strongly enriched in cases lacking the t(14;18), we selected for these otherwise rare subgroups among treatment-naive and relapsed FL and investigated the frequency and localization of NANGS, as well as IGHV gene usage.
Materials and methods
Study cohort
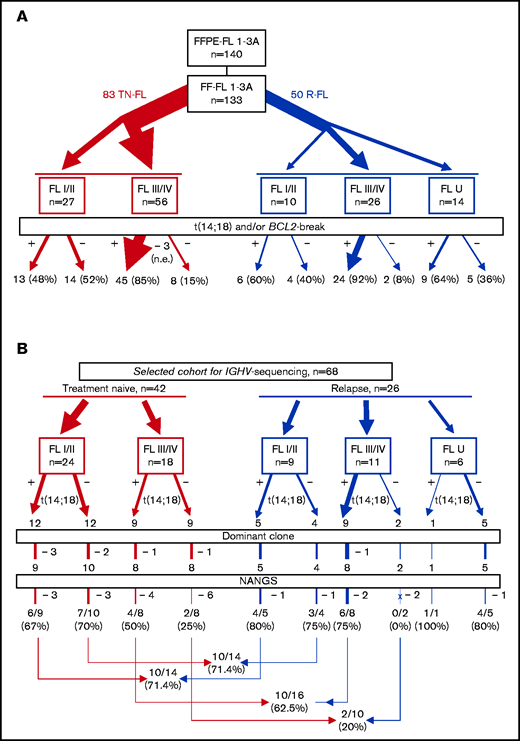
A total of 140 FL with formalin-fixed and paraffin-embedded material available (among them 133 with fresh frozen [FF] material with sufficient high-quality DNA accessible) were centrally reviewed, and a diagnosis of FL confirmed by 3 expert hematopathologists (A.R., G.O., A.Z.) (Figure 1A). Among the 133 FL with FF material, 83 were acquired at the time of diagnosis (referred to as treatment-naive FL), and 50 were sampled at first or subsequent relapse. Information on clinical stage could be obtained for 119 FL. Information on clinical stage at primary diagnosis was missing in a proportion of relapsed FL (10 FL stage I/II, 26 FL stage III/IV, 14 FL U [U = unknown clinical stage]). DNA from all t(14;18)-negative FL and a subset of t(14;18)-positive FL with FF material was used for sequencing (Table 1). Amplification and sequencing of the V-D-J sequences were performed in 12 t(14;18)-positive and 12 t(14;18)-negative treatment-naive FL stage I/II, 9 t(14;18)-positive and 9 t(14;18)-negative treatment-naive FL stage III/IV, and 15 t(14;18)-positive and 11 t(14;18)-negative relapsed FL stages I to IV.
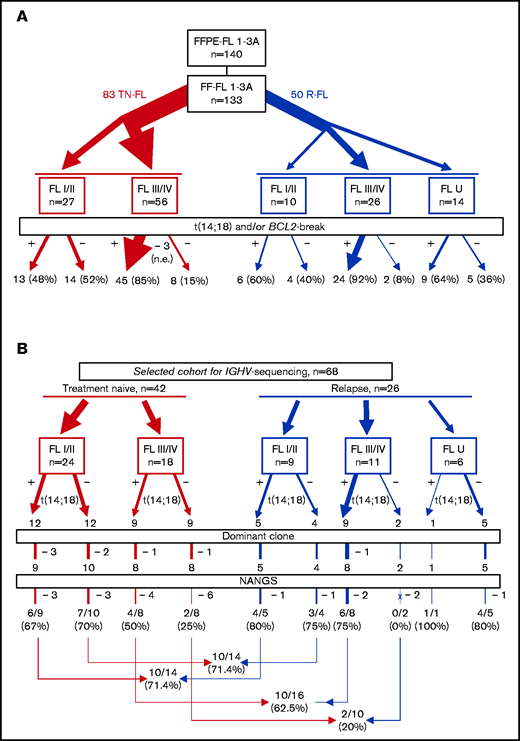
Flowchart depicting the study cohorts used for FISH analysis. (A) and IGHV-sequencing analysis (B) and the corresponding results. FFPE, formalin-fixed and paraffin-embedded; FF, fresh-frozen; I/II, FL at early clinical stage; III/IV, FL at advanced clinical stage; NANGS, newly acquired N-glycosylation sites; R, relapse; U, unknown clinical stage; TN, treatment-naive.
Flowchart depicting the study cohorts used for FISH analysis. (A) and IGHV-sequencing analysis (B) and the corresponding results. FFPE, formalin-fixed and paraffin-embedded; FF, fresh-frozen; I/II, FL at early clinical stage; III/IV, FL at advanced clinical stage; NANGS, newly acquired N-glycosylation sites; R, relapse; U, unknown clinical stage; TN, treatment-naive.
The study was approved by the local ethics committees of the participating and recruiting centers and was conducted in accordance with the Declaration of Helsinki.
Fluorescence in situ hybridization (FISH) analysis and immunohistochemistry
Clonotype and clonal frequency determination
For the detection of clonal V-D-J sequences of the IGHV gene, DNA was extracted from 10 to 15 10 µm slices of all FL with FF material available, using the AllPrep DNA Mini Kit (Qiagen GmbH, Hilden, Germany). Only samples with sufficient quality (260/280 ratio of ≥1.8) were included in the study. The V-D-J sequences were amplified by using the LymphoTrack Dx IGH FR1 Assay MiSeq kit (Invivoscribe, San Diego, CA), according to the manufacturer’s protocols. A leader-JH strategy (LymphoTrack IGHV Leader Somatic Hypermutation) was applied when primers did not bind to the FR1 region, probably due to an excess of SHM. Moreover, FR2 and FR3 strategies (LymphoTrack Dx IGH [FR1/FR2/FR3] Assays) were used when both strategies failed. If no clonal sequence was detected (at least 5% clonal frequency), a case was assigned as polyclonal.
Bioinformatics evaluation of sequencing data
To define the main clone, data were evaluated by using a self-developed pipeline based on pRESTO, Change-O, and Shazam,21-23 and the data output was subsequently compared with the results generated with the LymphoTrack software (MiSeq v2.1.1; LymphoTrack Software). In contrast to the LymphoTrack software, this pipeline includes all sequences in the analysis and accepts >2 different nucleotides given that the VH/JH is the same and CDR3 similar, which might be an advantage for tumors with high intraclonal diversification such as FL. Further details are provided in the supplemental Methods. Immunoglobulins were aligned against the IMGT reference database (http://www.imgt.org/IMGT_vquest/vquest) with IgBLAST (version 1.8.0). Moreover, the tools IMGT/V-Quest and IgBLAST (https://www.ncbi.nlm.nih.gov/igblast/) were used to manually analyze the sequences for NANGS (Asn-X-Ser/Thr, X can be any AA except Pro, Asp, or Glu).5 Sequencing data are deposited in the National Center for Biotechnology Information Sequence Read Archive database NCBI (accession number PRJNA722152). Sequences of the dominant clone are provided in supplemental Table 1. Notably, the 2 bioinformatics pipelines had a 100% concordance in terms of clonal detection, although not with regard to clonal frequency, probably due to the different algorithms mentioned earlier.
Statistical evaluation
Statistical data analysis was performed by using SPSS version 25 (IBM SPSS Statistics; IBM Corporation, Armonk, NY) and GraphPad Prism 8.3.1 (GraphPad Software, La Jolla, CA). Fisher’s exact test was applied to determine the significance (in general, two sided; one sided in case of a directed hypothesis) of the association between 2 variables. The Mann-Whitney U test was applied to compare 2 independent subgroups. Odds ratios (ORs) and 95% confidence intervals (CIs) were calculated with SciStat (OR calculator [scistat.com]). P values ≤.05 (Fisher’s exact test) were regarded as significant.
Results
Study cohort
In the whole data set of 133 FL with sufficient high-quality DNA available, ∼74% (96 of 130) of evaluable cases carried a BCL2-break (94 of 130 [∼72%] with verified IGH rearrangement), whereas 26% (34 of 130) of evaluable cases were both BCL2-break and t(14;18)-negative (Figure 1A). Because translocation partners for BCL2 other than IGH are rare in FL, and because there was a 100% concordance between the 2 FISH approaches concerning the cases of the sequencing cohort that could be successfully investigated with both approaches (66 of 68) (supplemental Table 2), we use the term t(14;18)-negative FL for all BCL2-break negative FL throughout this article. BCL2 expression in >30% of tumor cells was detected in 97% of t(14;18)-positive and 82% of t(14;18)-negative FL. Treatment-naive FL stage I/II showed BCL2-breaks in 13 (48%) of 27 cases, whereas 14 (52%) of 30 were t(14;18) negative. In contrast, the vast majority of evaluable treatment-naive FL stage III/IV (45 of 54 [83%]) were t(14;18)-positive, whereas only a minority of FL stage III/IV were t(14;18)-negative (9 of 54 [17%]). Among 50 relapsed FL, 10 were FL stage I/II [t(14;18)-positive in 6 of 10 (60%) cases] and 26 FL stage III/IV [24 of 26 (92%) t(14;18)-positive]. Notably, 5 (36%) of 14 relapsed FL U were t(14;18)-negative. No transformed FL cases were included in the study.
Clonotype and clonal frequency detection
Sequencing of the IGHV regions was performed by using FF material of 68 FL, namely all 32 t(14;18)-negative FL and 36 t(14;18)-positive FL with high-quality DNA available (Figure 1B; Table 1). Using the pipeline based on pRESTO, Change-O and Shazam could determine the main clone (>5%) and a productive clonal sequence in 60 (~88%) of 68 samples; in 8 of 68 FL (∼12%), we could not detect a productive clone with >5% frequency according to any approach (Figure 1B; supplemental Table 2). The median clonal frequency among all cases was 61.2%. Three cases had a clonal frequency of <10% by using the FR1- or the leader-JH approach; however, a clonal frequency of >50% of the same clone was revealed by using either the FR2 or the FR3 approach (supplemental Table 3). While FL stage I/II (including relapsed FL) had a median clonal frequency of 51.2%, the median clonal frequency of FL stage III/IV (including relapsed FL) was 75.4% (P = .015) (supplemental Table 4; supplemental Figure 1A). Instead, no significant differences were observed in the clonal frequency between t(14;18)-positive and t(14;18)-negative FL, nor between treatment-naive and relapsed FL (supplemental Table 4; supplemental Figure 1B-C). Notably, the LymphoTrack software confirmed the identified clones, however, revealed a different clonal frequency. The median SHM rate ranged from 9.1% to 16.6% among the different subgroups; however, the SHM rate did not differ significantly between treatment-naive and relapsed FL, FL stage I/II and FL stage III/IV, and t(14;18)-positive and t(14;18)-negative FL, respectively.
NANGS are significantly reduced in high advanced-stage but not in low early-stage t(14;18)-negative FL
In a previous study with few t(14;18)-negative FL of an unselected cohort, we reported that NANGS are less common in t(14;18)-negative FL compared with “classical” t(14;18)-positive FL.18 Thus, we aimed to validate these findings in an independent cohort and to compare early- and advanced-stage FL for the occurrence of NANGS. The overall frequency of NANGS in the entire current study cohort was 61.7% (20 of 28 [71.4%] for FL stage I/II [treatment-naive plus relapsed FL], 12 of 26 [46.2%] for FL stage III/IV [treatment-naive plus relapsed FL], and 83.3% for relapsed FL U) (Figure 1B). Including both treatment-naive and relapsed FL, the number of NANGS differed significantly between t(14;18)-positive and t(14;18)-negative FL stage III/IV (10 of 16 vs 2 of 10; P = .042 [one-sided Fisher’s exact test, directed hypothesis]; OR, 6.66; 95% CI, 1.04-42.43; z-statistic, 2.0 [associated P value = .044) (Table 2; Figure 1B), whereas no significant difference could be shown between t(14;18)-positive and t(14;18)-negative FL stage I/II (10 of 14 vs 10 of 14). Consequently, a significant difference was observed between t(14;18)-negative FL stage I/II and t(14;18)-negative FL stage III/IV, whereas no significant difference was observed between t(14;18)-positive FL stage I/II and t(14;18)-positive FL stage III/IV. In summary, advanced-stage, t(14;18)-negative FL display a lower NANGS content compared with any other group [ie, early-stage t(14;18)-negative and t(14;18)-positive in either group].
Localization and hotspots of NANGS
Because previous studies in unselected cohorts of FL showed a predominant localization of NANGS in CDR2,5 we aimed to assess the localization of NANGS in our study cohort that was enriched for the rare subgroups of t(14;18)-negative FL and early-stage I/II FL. Considering the whole study cohort, NANGS mainly clustered in CDR3 and FR3. Specifically, 17 (34%) of 50 NANGS were located in CDR3 and 13 (26%) of 50 in FR3 (Figure 2A; supplemental Table 2). The remaining sites were distributed to CDR1/FR2 (7 of 50 [14%]), FR2/CDR2 (6 of 50 [12%]), CDR2 (4 of 50 [8%]), CDR1 (1 of 50 [2%]), CDR3/FR4 (1 of 50 [2%]), and FR4 (1 of 50 [2%]). Hotspots at high frequency were found in CDR1/FR2 (7 of 7 at AA 38-40, corresponding to the AA 33-35 in CDR1 according to Kabat numberings24 ) and FR2/CDR2 (6 of 6 at AA 55-57, equivalent to the AA 50-52 in CDR2 according to Kabat numberings) and hotspots at lower frequency in CDR3 (8 of 17 at AA 107-109), CDR2 (2 of 4 at AA 63-65), and FR3 (4 of 14 at AA 66-68, 3 of 14 at AA 68-70, and 3 of 14 at AA 90-92) (supplemental Figure 2).
![Local distribution of newly acquired N-glycosylation sites (NANGS) in the entire cohort (FL stage I-IV, including relapsed FL). (A), in t(14;18)-positive [t(14;18) +] compared with t(14;18)-negative [t(14;18) -] FL of the entire cohort (B), and in t(14;18) + and t(14;18) - FL stages I to IV at relapse (R-FL I-IV), including FL of unknown stage (R-FL U) (C). FL I/II, early-stage FL; FL III/IV, advanced-stage FL.](https://ash.silverchair-cdn.com/ash/content_public/journal/bloodadvances/5/23/10.1182_bloodadvances.2021005081/2/m_advancesadv2021005081f2.png?Expires=1765081268&Signature=uQD0M4484zIXjAr7YFM~hSjAU~JB0A-ZXO-~KccwEQChTjRVjOg~x5OvPhgluR31MQ9Xu6JK429WiE6ltrnftQnxQQ~kTVOSYI07J1Bhc5V-PMv16ogits4cQFSAay44x-LW5R1iV-LQ9H7lpDDnSmNO8JlPnfXHZ7RYl~Z5UDRnEtqsvS6SbyFfrDdQjwr-lws33As49mNhZBifXdrmD8ZCX-6y23BU7Psl5pBw6xIdJLonL1hrnHxS~3piMgTrcnftZxERdkLbxlCUiePhvNwRBvZkb8~oCe5myZS0sNAUvDPJvW8jmtA5qyDklSKg6ZS2MOsvog0kjVnB~jFNtw__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Local distribution of newly acquired N-glycosylation sites (NANGS) in the entire cohort (FL stage I-IV, including relapsed FL). (A), in t(14;18)-positive [t(14;18) +] compared with t(14;18)-negative [t(14;18) -] FL of the entire cohort (B), and in t(14;18) + and t(14;18) - FL stages I to IV at relapse (R-FL I-IV), including FL of unknown stage (R-FL U) (C). FL I/II, early-stage FL; FL III/IV, advanced-stage FL.
Local distribution of newly acquired N-glycosylation sites (NANGS) in the entire cohort (FL stage I-IV, including relapsed FL). (A), in t(14;18)-positive [t(14;18) +] compared with t(14;18)-negative [t(14;18) -] FL of the entire cohort (B), and in t(14;18) + and t(14;18) - FL stages I to IV at relapse (R-FL I-IV), including FL of unknown stage (R-FL U) (C). FL I/II, early-stage FL; FL III/IV, advanced-stage FL.
t(14;18)-negative FL display a strong enrichment of NANGS in FR3
Irrespective of clinical stage and status at biopsy, t(14;18)-negative FL showed a striking enrichment of NANGS in the FR3 region compared with t(14;18)-positive FL (12 of 24 [50%] vs 1 of 26 [∼4%]; P < .001; OR, 0.04; 95% CI, 0.0046-0.34) (Figure 2B; supplemental Figure 3). Notably, the mutation hotspots at AA 68-70 and 90-92 in FR3 were restricted to t(14;18)-negative FL. Compared with t(14;18)-positive FL, t(14;18)-negative FL also showed a lower frequency of NANGS in the regions of CDR1/FR2 (1 of 24 [4%] vs 6 of 26 [23%]; not significant; OR, 6.9; 95% CI, 0.76-62.28), FR2/CDR2 (0 of 24 [0%] vs 6 of 26 [23%]; P = .02; OR, 15.53; 95% CI, 0.82-292.63), and CDR3 (6 of 24 [25%] vs 11 of 26 [42%]; P = .2; OR, 2.2; 95% CI, 0.65-7.36). The region that was reported by Zhu et al5 to be most frequently affected (CDR2, here: FR2/CDR2) was mainly affected in t(14;18)-positive FL at relapse (4 of 6 cases with NANGS in FR2/CDR2) (Figure 2C; supplemental Table 2).
Differences between FL with and without t(14;18) in the generation of NANGS
Because the probability of being glycosylated might be higher or lower depending on the specific AA composition of the N-glycosylation motive,25 we investigated the precise AA sequences of NANGS in the current study cohort. Twenty-one different AA motives were detected in 37 FL samples with NANGS. The most recurrent motives were NIS (n = 8), NMT (n = 4), NLS (n = 4), NIT (n = 4), and NYT (n = 4). The AA Asn/N at position one was introduced by mutations in all (24 of 24) t(14;18)-positive FL during the generation of NANGS, whereas it was already present in 8 of 23 NANGS (∼35%) of t(14;18)-negative FL (P = .02; OR, 89.3; 95% CI, 4.80-1660.52) (supplemental Figure 2). Moreover, the motive NLS was detected selectively in four t(14;18)-negative FL, predominantly in FL stage I/II (3 FL stage I/II, 1 FL stage III/IV). Finally, of all NANGS in CDR1-FR3 with a threonine at position 3 (n = 15), 8 threonines were derived by a mutation from serine: specifically, 6 of 8 (75%) in five t(14;18)-positive FL and 2 of 8 (25%) in two t(14;18)-negative FL with an equal distribution among clinical stages (3 FL stage I/II, 2 FL stage I/II, and 2 FL U) (supplemental Figure 2).
IGHV4-34 usage in FL with NANGS is restricted to t(14;18)-negative treatment-naive FL
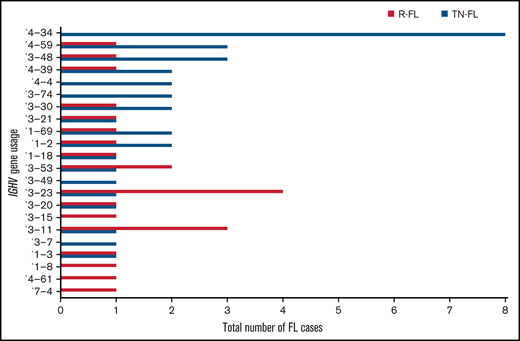
Because a biased practice of IGHV usage has been described in many lymphoma entities such as diffuse large B-cell lymphoma (DLBCL) and chronic lymphocytic leukemia (CLL), and might influence the biology of the neoplastic clone,26-31 we compared t(14;18)-positive and t(14;18)-negative FL, as well as different clinical subgroups of the current cohort, in relation to this feature. We found a preference for IGHV4-34 usage in treatment-naive FL, whereas it was completely absent in relapsed FL (8 of 35 treatment-naive vs 0 of 25 relapsed; P = .016; OR, 15.76; 95% CI, 0.86-287.25) (Figure 3). Moreover, IGHV4-34 usage was found more often in t(14;18)-negative FL compared with t(14;18)-positive FL of the whole sequenced cohort with clonal amplification (6 of 29 [∼21%] vs 2 of 31 [∼6%]; P = .14; OR, 3.8; 95% CI, 0.69-20.52) and with restriction to treatment-naive FL (6 of 18 [∼33%] vs 2 of 17 [12%]; P = .23; OR, 3.75; 95% CI, 0.63-22.04) (Table 3). Notably, focusing on FL with NANGS, 5 of 9 (55.5%) t(14;18)-negative FL (only treatment-naive) used the IGHV4-34 gene, whereas it was completely absent in 10 t(14;18)-positive FL (5 of 9 vs 0 of 10; P = .01; OR, 25.66; 95% CI, 1.15-568.94). Notably, the D- and J-segments varied among FL with IGHV4-34 usage (supplemental Figure 1).
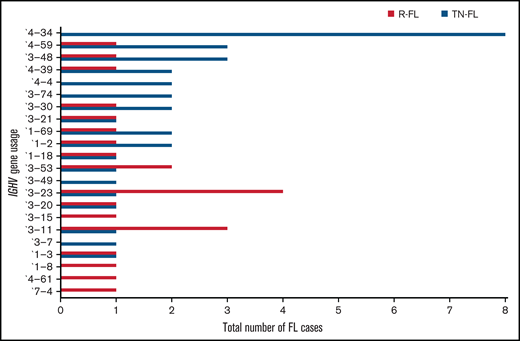
IGHV gene usage in treatment-naive (TN) and relapsed (R) FL. The figure shows both treatment-naive and relapsed FL.
IGHV gene usage in treatment-naive (TN) and relapsed (R) FL. The figure shows both treatment-naive and relapsed FL.
Discussion
FL is a GC-derived B-cell lymphoma that, despite the occurrence of the t(14;18) in most cases, expresses an intact functional BCR.5 FL in unselected cohorts [predominant t(14;18)-positive advanced stage III/IV FL] have been shown to harbor a high frequency of NANGS in their immunoglobulin chains that allows the tumor cells to interact with the microenvironment via lectins expressed by M2 macrophages.16 Notably, the frequency of NANGS in FL is higher compared with most other GC-derived malignancies and specifically higher than in post-GC B cell–derived malignancies.5,32 Moreover, it was shown that these NANGS are functionally active, in contrast to naturally occurring sites,16 and they are retained in relapse and transformation events,7 suggesting that NANGS are relevant in the pathogenesis of FL. However, thus far, little is known about NANGS in t(14;18)-negative and stage I/II FL.
To gain more knowledge about the presence and localization of NANGS in those rare subgroups at primary diagnosis and at relapse, we enriched our FL cohort for these cases and compared their features with the more prevalent t(14;18)-positive and stage III/IV subgroups.
Although previous studies in unselected cohorts revealed NANGS in the IGHV genes of 80% to 100% of FL,5,6,8 we detected those sites in only 61.7% of cases in our entire cohort, which seems to be predominantly a consequence of case selection in the current cohort.
In line with previous findings,18 NANGS were absent in the vast majority of t(14;18)-negative FL stage III/IV cases, while present in the majority of t(14;18)-positive FL.18,32-34 Of note, the frequency of SHM in t(14;18)-positive and t(14;18)-negative FL stage III/IV was comparable and thus does not account for the reduced number of NANGS in t(14;18)-negative FL stage III/IV. However, the AA sequence NLS, which is less prone to N-glycosylation, was selectively found in t(14;18)-negative FL,25 and the replacement of serine at position 3 of the AA sequence, increasing the probability of an N-glycosylation,25 was more often found in t(14;18)-positive FL.
In contrast to FL stage III/IV, the majority of the IGHV genes of both t(14;18)-negative and t(14;18)-positive FL I/II harbored NANGS at comparable frequencies. Thus, t(14;18)-negative FL stage III/IV exhibit significantly lower levels of NANGS than t(14;18)-negative FL stage I/II. Together with the finding of a significantly lower clonal frequency in FL stage I/II compared with FL stage III/IV, these observations suggest not only differences in the pathogenesis of t(14;18)-positive and t(14;18)-negative FL stage III/IV but also between early- and advanced-stage FL.
Moreover, differences in localization of the mutated sites and the IGHV usage were observed among the different FL subgroups. An enrichment of NANGS in CDR2, the most common site in previously published FL cohorts,5 was mainly found in t(14;18)-positive relapsed FL stage I to IV in our study, which is the subgroup that probably most closely matches the previously investigated cohorts. Notably, the N-glycosylation hotspots we found were the same reported in previous publications using the Kabat nomenclature,24 although apparently at different positions due to the various nomenclatures used.
At variance with previous findings, t(14;18)-negative FL stages I/II and III/IV were strongly enriched in NANGS in FR3. Interestingly, N-glycosylation in this region has been associated with different autoimmune diseases such as rheumatoid arthritis and Sjögren’s syndrome,32 suggesting that t(14;18)-negative FL use a mechanism that is possibly associated with autoreactivity. However, although the variable domain glycans in autoimmune diseases are highly sialylated, they are rich in high-mannose structures in FL patients of unselected cohorts.32,35 This leads to the assumption that the type of N-glycosylation might also differ between t(14;18)-positive and t(14;18)-negative FL. This hypothesis, however, requires detailed functional investigations that go beyond the scope of the current work.
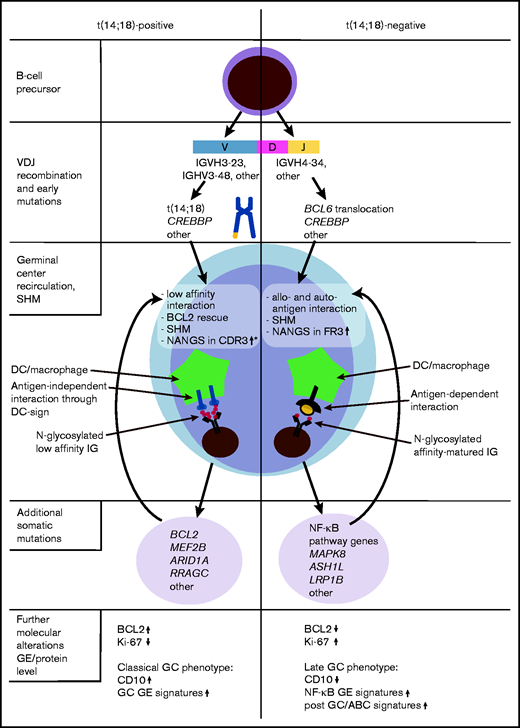
The theory of enhanced autoreactivity in t(14;18)-negative FL is furthermore supported by the enriched IGHV4-34 usage, which was specifically prominent in t(14;18)-negative treatment-naive FL with NANGS (Figure 4). Notably, the IGHV4-34 gene, also known as IGHV4-21, alone (without stereotyped associations) is linked to autoimmunity and can produce autoantibodies such as cold agglutinins, rheumatoid factor, and anti-DNA antibodies that bind to structurally distinct auto- and allogeneic antigens.37-40,41 Interestingly, IGHV4-34 usage was rare in t(14;18)-positive FL (only natural sites without NANGS) and absent in treatment-naive cases of t(14;18)-positive FL stage I to IV with NANGS. Intriguingly, IGHV4-34 usage was absent in relapsed cases from our cohort. This finding could suggest that cases with IGHV4-34 usage may tend to transform to a more aggressive disease rather than relapsing as FL, a hypothesis supported by the presence of this IGHV gene in a proportion of DLBCL and transformed FL.29,31,42 If independently confirmed in larger cohorts, this feature might help to improve the molecular stratification of FL.
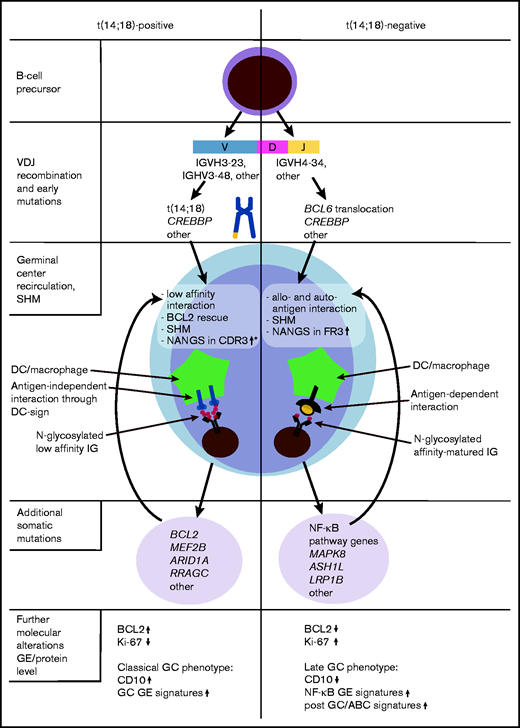
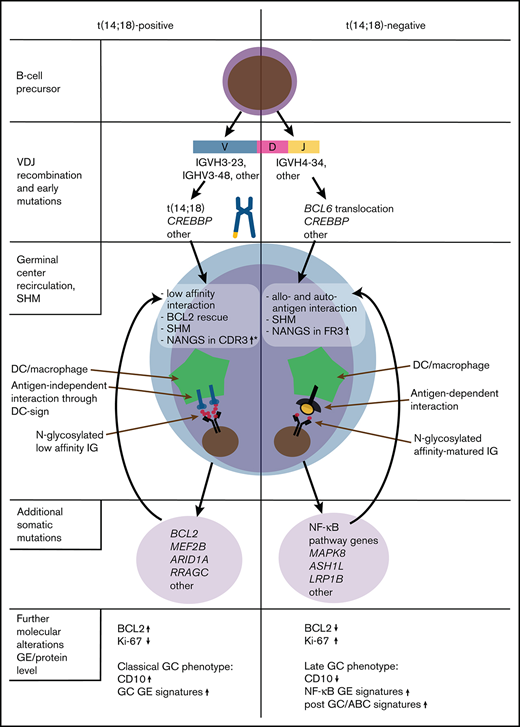
Hypothesis-driven model depicting the molecular pathogenesis of t(14;18)-positive and t(14;18)-negative FL, according to current and previous findings.3,18,33,34 *Gains of newly acquired N-glycosylation sites (NANGS) in CDR3 were specifically prominent in t(14;18)-positive treatment-naive FL stage I/II. DC, dendritic cell; GE, gene expression.
Hypothesis-driven model depicting the molecular pathogenesis of t(14;18)-positive and t(14;18)-negative FL, according to current and previous findings.3,18,33,34 *Gains of newly acquired N-glycosylation sites (NANGS) in CDR3 were specifically prominent in t(14;18)-positive treatment-naive FL stage I/II. DC, dendritic cell; GE, gene expression.
The natural NANGS found in IGHV4-34 in CDR2 were lost in 50% of cases and thus did not seem to play a specific role in our cohort. This finding confirms previous results, showing that these sites are not oligo-mannosylated and therefore not capable of interacting with lectins.8
IGHV4-34 usage is a specific feature of activated B-cell like (ABC) DLBCL and mutated CLL.8,29,31,43 Interestingly, several of our previous works showed that a subset of t(14;18)-negative FL have a “late” or “exit” GC phenotype,3,18,33,34 resembling the molecular phenotype described for ABC-DLBCL (Figure 4). In support of these findings, t(14;18)-negative FL were found to preferentially transform into ABC-DLBCL.44 Moreover, it was shown that ABC-DLBCL cells with IGHV4-34 usage reacted with autoantigens,31 which supports our hypothesis that a subgroup of t(14;18)-negative FL might also be stimulated by autoantigens. Overall, these molecular similarities may suggest that at least a subgroup of t(14;18)-negative FL might be targeted by a special therapeutic approach, in part reflecting what was proposed for molecular DLBCL subgroups.45 However, because classical FL is an indolent tumor that needs its microenvironment for growth and survival,2 no classical FL cell lines exist, and in vitro cultivation of primary FL cells, as well as the establishment of in vivo FL models, is a great challenge.46,47 Investigations are either limited to transformed FL or t(14;18)-positive, aggressive B-cell non-Hodgkin lymphoma cell lines48,49 or to a few in vitro or in vivo models that might be representative for classical indolent FL.50,51 Thus, it is currently almost impossible to perform functional studies that could provide the preclinical rationale to design approaches specific for t(14;18)-negative FL in the context of clinical trials.
In summary, we observed that NANGS obviously are less common in t(14;18)-negative advanced-stage (III/IV) FL, whereas no reduction was observed in t(14;18)-negative early-stage (I/II) FL (Figure 4). A significant part of t(14;18)-negative FL stage III/IV thus might take advantage of antigen-dependent BCR stimulation, which may provide alternative survival and proliferation signals to the antigen-independent, lectin-mediated BCR activation described in classical FL. Moreover, the high IGHV4-34 usage in treatment-naive t(14;18)-negative FL with NANGS and the strong accumulation of NANGS in FR3 might be an indication of the responsiveness to autoantigens in this FL subgroup.
A limitation of our study is the relatively low number of cases analyzed, which is a consequence of the rare occurrence of t(14;18)-negative and early-stage FL and the lack of FF tissue for most of the archived samples. Our findings therefore need to be confirmed in independent studies.
Nevertheless, the results of the current study might be of value to understand the clinical and molecular complexity of the different FL subgroups, which might partly be explained through the understanding of its complex immunogenetics. Resolving this complexity may ultimately provide a rationale for better risk stratification and the design of more personalized therapies in the context of clinical trials.
Acknowledgments
The authors kindly thank Theodora Nedeva and Tina Grieb for their excellent technical assistance, Alexander Kerscher for the clinical database support, and Bernhard Puppe for statistical support.
H.H. and G.O. are supported by the Robert-Bosch-Stiftung (Project Grant O3). V.G. is supported by Progetto Ricerca Finalizzata PE-2016-02362756 and RF-2018-12365790, Italian Ministry of Health; the Associazione Italiana Ricerca Cancro, Investigator Grant IG-21687; and the Associazione Italiana contro le Leucemie, linfomi e mielomi (AIL), Venezia Section, Pramaggiore/Veneto Orientale Group.
Authorship
Contribution: E.L., C.M., R.B., and F.V. performed research; E.L. and A.Z. conceived and designed the research study; E.L., C.M., R.B., F.V., A.B., H.H., V.G., and G.O. acquired data; E.L. and A.Z. supervised the study; E.L., C.M., R.B., F.V., A.R., and A.Z. analyzed and interpreted the data; E.L, C.M., and A.Z. drafted the manuscript; E.L., C.M., R.B., H.H., G.O., A.R., and A.Z. revised the manuscript; and all authors approved the final version and will ensure that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Ellen Leich-Zbat, Institute of Pathology, University of Würzburg, Josef-Schneider-Str. 2, 97080 Würzburg, Germany; e-mail: ellen.leich@uni-wuerzburg.de.

![Local distribution of newly acquired N-glycosylation sites (NANGS) in the entire cohort (FL stage I-IV, including relapsed FL). (A), in t(14;18)-positive [t(14;18) +] compared with t(14;18)-negative [t(14;18) -] FL of the entire cohort (B), and in t(14;18) + and t(14;18) - FL stages I to IV at relapse (R-FL I-IV), including FL of unknown stage (R-FL U) (C). FL I/II, early-stage FL; FL III/IV, advanced-stage FL.](https://ash.silverchair-cdn.com/ash/content_public/journal/bloodadvances/5/23/10.1182_bloodadvances.2021005081/2/m_advancesadv2021005081f2.png?Expires=1765591832&Signature=ml~4v-V6ZPtdc16OjlZW8jDezabOx0rLS0zD0Y6PuFUxTEUznutjx5Tin1clfs897DPRyP7IQgGj-A3HMZa1ymzbaP2cjtTJVeA9l2xYK8lVsOKC81dwQMYQlotxoutcns8DR7EaFCtpSz5JGIbkZoG08x7sGxhRFTT-LTEsMlXyNi-EM8cysuULjRiiXaZbNURKrScJKGH0p3VaQOORgILCRod6sQM7aoll6yeNjwgm2d8PvcsvIAnEEusj0d7ze5HTJm8ZQW5h0hvBFyXt8ygXcYejHi1tBvEKI0B4MtHI5eZ3i6i0eFL6VyGbeM7YHFD~-j5FtJwVtDquB~kbeA__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)