

Checkpoint blockade has become a promising immunotherapy for the treatment of a variety of malignancies. In particular, the receptor programmed death-1 (PD-1) has become a focus of intense study due to its expression on and negative regulation of T-cell function. The ligand for PD-1, PD-L1, is upregulated on many tumors and, as a result, can suppress antigen-specific T-cells thereby limiting their anti-tumor response. Pharmacological PD-1/PD-L1 axis disruption can occur with either Pembrolizumab and Nivolumab (PD-1 antagonists) and Avelumab and Atezolizumab (PD-L1 antagonists). These antibodies (mAbs) are being used to treat melanoma, non-small cell lung cancer, kidney, bladder and head and neck cancer with varying degrees of success. Like T-cells, natural killer cells (NK) also have potent antitumor cytolytic properties. The expression and functional effects of PD-1 on NK cells remain unclear due to difficulties in receptor detection and efficacy of receptor blockade by available commercial reagents. While some studies have been unable to detect PD-1 on resting NK cells, others have identified PD-1 expression only on specific NK populations under certain conditions (e.g. Cytokine stimulation or virus infection).
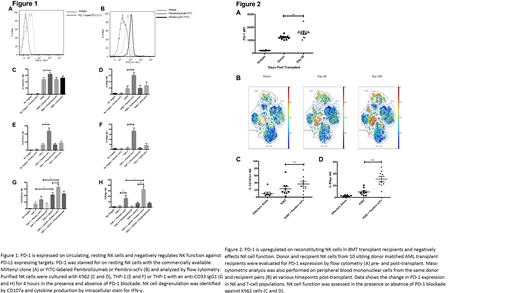
Here, we identify PD-1 expression on peripheral blood NK cells. Using commercial reagents (Figure 1A) and a FITC-labeled clinical mAb (Pembrolizumab, Pembro), we detect low yet consistent PD-1 expression on all circulating, resting NK cells. Since FITC-Pembro mean fluorescent intensity was low and a high proportion of FITC labeled NK cells overlapped with the isotype control (Figure 1B), we designed a short-chain variable fragment (scFv) of the mAb to determine whether the smaller scFv molecule has better binding and functional activity than the intact mAb. The Pembro scFv bound to resting NK cells with a distinct fluorescent peak compared to the native Prembro from which the scFv was derived (Figure 1B). Compared to intact Prembro, use of the Pembro scFv as a PD-1 antagonist resulted in a 2-fold increase of NK cell cytolytic activity and a 3-4 fold increase in cytokine production against the PD-L1 expressing CML target, K562 (Figure 1C-D) and the AML target, THP-1 (Figure 1E-F). While PD-1 blockade enhanced NK cell degranulation and target cell killing, a greater functional enhancement was seen for interferon-γ production. PD-1 signaling inhibits PI3K induced pAkt and NK function. PD-1/PD-1 ligand blockade by the Pembro scFv resulted in increased NK cell pAKT in the presence of PD-L1 and NK activating NKG2D-ligand-expressing THP-1 cells. In addition to natural cytotoxicity, NK-mediated ADCC was also enhanced with PD-1 blockade. CD33 mAb immunoconjugates have been used to treat AML. Combined anti-CD33 mAb and PD-1 blockade against THP-1 cells resulted in a small but significant increase in NK cell degranulation and a 4-fold increase in cytokine production compared to anti-CD33 mAb without PD-1 blockade (Figure 1G-H). Since stimulation with IL-15, a cytokine that effectively lowers the NK activation threshold, abrogated the benefits of Pembro scFv in diminishing PD-1 inhibitory effects on NK cells, PD-1 control of NK function appears limited to be mostly relevant to resting NK cells. To understand the physiologic expression of PD-1 in vivo, we studied samples taken from AML patients receiving matched sibling donor transplantation at the University of Minnesota. Increased PD-1 on reconstituting NK cells in BMT recipients up to day 100 post-transplant was shown by both flow-cytometric (Figure 2A) and mass-cytometric (CyTOF) analyses (Figure 2B). Blockade of PD-1 on these cells significantly enhanced both NK degranulation (Figure 2C) and cytokine production (Figure 2D) against K562 targets. A similar increase in NK function was observed with PD-1 blockade in AML patients receiving umbilical cord transplants (not shown). These data indicate that PD-1 is present on human NK cells and PD-1 ligation negatively regulates NK function against PD-L1 expressing tumor targets. The observation that functional PD-1 is expressed on NK cells under resting conditions strongly suggests that the use of a PD-1 antagonist, in combination with NK cell therapy, should be clinically effective for treatment of cancer.
Felices:GT Biopharma.: Other: consulting funds, Research Funding. Blazar:Kamon Pharmaceuticals, Inc: Membership on an entity's Board of Directors or advisory committees; Tmunity: Other: Co-Founder; BlueRock Therapeutics: Membership on an entity's Board of Directors or advisory committees; Regeneron Pharmaceuticals: Membership on an entity's Board of Directors or advisory committees; Five Prime Therapeutics Inc: Co-Founder, Membership on an entity's Board of Directors or advisory committees; KidsFirst Fund: Research Funding; Childrens' Cancer Research Fund: Research Funding; Leukemia and Lymphoma Society: Research Funding; Abbvie Inc: Research Funding; Alpine Immune Sciences, Inc.: Research Funding; RXi Pharmaceuticals: Research Funding; Fate Therapeutics, Inc.: Research Funding; Magenta Therapeutics and BlueRock Therapeuetics: Membership on an entity's Board of Directors or advisory committees. Vallera:GT Biopharma, Inc.: Consultancy, Research Funding. Miller:Fate Therapeutics, Inc: Consultancy, Research Funding; GT BioPharma: Consultancy, Membership on an entity's Board of Directors or advisory committees, Research Funding; CytoSen: Membership on an entity's Board of Directors or advisory committees; OnKImmune: Membership on an entity's Board of Directors or advisory committees; Dr. Reddys Laboratory: Membership on an entity's Board of Directors or advisory committees; Moderna: Membership on an entity's Board of Directors or advisory committees.
Keytruda. PD-1 blockade on NK cells for tumor immunotherapy

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal