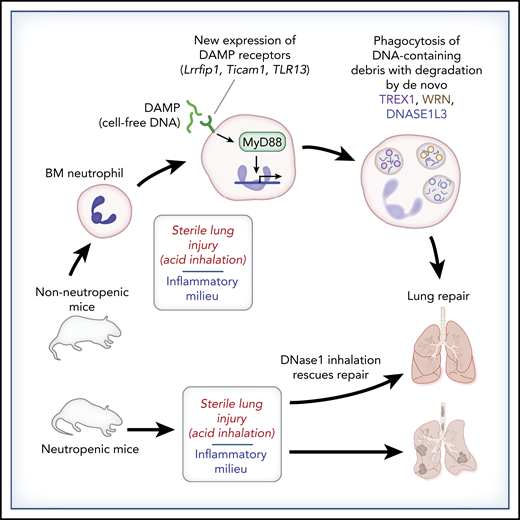
In this issue of Blood, 1 further debunk the anachronistic notion that neutrophils are merely vehicles for presynthesized and prepackaged antimicrobial factors and degradative enzymes. Their report describes a nuanced role for neutrophils in recovery from experimental sterile acute lung injury through new gene expression that enables clearance of DNA-containing debris from areas of lung injury (see figure).
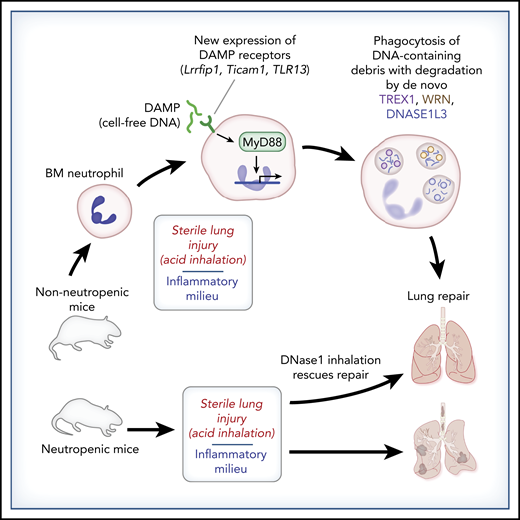
Neutrophil clean-up in experimental acute sterile lung injury: de novo DNA degradation leads to lung repair. Bone marrow–derived neutrophils (BM neutrophils) lack the ability to degrade DNA-containing debris. However, alveolar neutrophils acquire this ability following exposure to an inflammatory milieu. Oved et al demonstrate that sterile acute lung injury following acid inhalation induces de novo expression of DNA-sensing DAMP receptors, MyD88-dependent transcription, and protein synthesis of DNA-degradative factors (TREX1, WRN, DNASE1L3), leading to clearance of DNA-containing debris that otherwise hinders lung recovery. Neutropenic mice do not clear DNA-containing lung debris in this model, leading to failed lung repair, but tracheal treatment with DNase I rescues the lung repair phenotype.
Neutrophil clean-up in experimental acute sterile lung injury: de novo DNA degradation leads to lung repair. Bone marrow–derived neutrophils (BM neutrophils) lack the ability to degrade DNA-containing debris. However, alveolar neutrophils acquire this ability following exposure to an inflammatory milieu. Oved et al demonstrate that sterile acute lung injury following acid inhalation induces de novo expression of DNA-sensing DAMP receptors, MyD88-dependent transcription, and protein synthesis of DNA-degradative factors (TREX1, WRN, DNASE1L3), leading to clearance of DNA-containing debris that otherwise hinders lung recovery. Neutropenic mice do not clear DNA-containing lung debris in this model, leading to failed lung repair, but tracheal treatment with DNase I rescues the lung repair phenotype.
Neutrophils are routinely implicated in the tissue damage associated with uncontrolled and dysregulated pulmonary inflammation.2 As the first cellular responders to infection or tissue damage, neutrophils often play the role of primary effector cell in an all-out response to injury, come what may. The collateral tissue damage released following delivery of such a robust payload by neutrophils is well described and has led many investigators to consider the neutrophil as a target for anti-inflammatory drug development.3 The discovery and characterization of neutrophil extracellular trap (NET) formation with its potential to participate pathogenically in autoimmune and inflammatory syndromes, such as systemic lupus erythematosus, sepsis, atherosclerosis, and most recently, COVID-19, highlights the potential for DNA- and chromatin-mediated inflammatory tissue damage in infectious and sterile milieus.4,5 Indeed, past characterizations of neutrophil function in the response to sterile tissue injury are particularly damning.
The combined work by Oved et al and other investigators6,7 will change that perception. In their Brief Report, Oved et al report on the impactful and particularly striking finding of differential DNA-degradative capacity between neutrophils leaving the bone marrow and bronchial alveolar neutrophils recruited to the site of sterile injury in this model. They elegantly demonstrate induction of DNA-sensing damage-associated molecular pattern (DAMP)-recognition molecules (Lrrfip1, Ticam1, and TLR13) in neutrophils exposed to the sterile inflammatory milieu. Furthermore, subsequent DAMP receptor– and MyD88-dependent signaling enables the neutrophil to respond to sterile acute lung injury in a reparative manner, with de novo transcription and translation of the DNA-degradative factors TREX1, WRN, and DNASE1L3 occurring in response to the inflammatory milieu encountered by migrating neutrophils. Thus, through de novo induction of DAMP receptors, resultant MyD88-dependent transcription, and translation of DNA-degrading proteins, neutrophils are “finished and prepared” for their reparative role in this model of sterile acute lung injury.
This Brief Report also highlights the impact of DNA-containing debris in delaying lung recovery in this model of sterile acute lung injury. They found that neutropenic mice fail to clear chromatin-containing inflammatory debris following acid inhalation. Intratracheal treatment with recombinant DNase I rescues the reparative phenotype, suggesting that cell-free DNA and chromatin potentiate and extend inflammation at sites of tissue injury and delay healing. Although the mechanisms through which cell-free DNA and chromatin hinder tissue repair extend beyond the scope of this Brief Report, histone-complexed DNA fragments do correlate with endothelial cell damage, coagulopathy, and mortality in pediatric trauma patients.8
Finally, an interesting question raised by this study is whether non-NETing neutrophils in the inflammatory milieu, sterile or otherwise, acquire similar capabilities to aid in the clearance of DNA-containing debris from ongoing NET formation by a subset of neutrophils recruited to the area of injury. With the link between failed clearance of self-DNA debris and autoimmune disease clearly demonstrated,9 failure of neutrophil-mediated self-DNA debris clearance, as demonstrated in this model, could contribute to an increased risk for autoimmune disease in patients with inflammatory syndromes and NET formation. It will be interesting to see whether non-NETing neutrophils are “prepared and finished” by the inflammatory milieu to clear the NETotic debris.
Conflict-of-interest disclosure: The author declares no competing financial interests.