

Key Points
3D organoid T-cell differentiation from a few hundred peripheral blood CD34+ cells was successfully achieved.
3D organoid T-cell differentiation could help physicians distinguish intrinsic from extrinsic defects underlying a clinical SCID phenotype.

Introduction
Newborn screening in North America has led to an increased incidence of patients diagnosed with severe combined immune deficiency (SCID).1 Although many SCID-causing genes have been identified, clinicians may face a patient without any abnormal gene identified even with advanced sequencing technologies. In a recent SCID prospective cohort, 7% of patients did not present any genetic variants that could be ascribed to known SCID-causing genes.2 From the perspective of hematopoietic stem cell (HSC) differentiation, the mechanism of the disease could be intrinsic, affecting their differentiation capacity, or/and extrinsic, depending on outside instructive cues (ie, the thymus). There is a dire need of a test that would guide the physician to discriminate between bone marrow (BM) and thymus transplantation for unclassified SCID patients. Considering this unmet clinical need, our objective was to set up a functional test that would be feasible using a small volume of peripheral blood (PB).
Methods
The study was approved by CHU Sainte-Justine institutional review board (protocols 3195 and 2126). Samples were obtained after obtaining informed consent from patients and institutional review board–approved biobanks. A summary of patients is shown in supplemental Table 1.
OP9-DLL4 cells were generated by retroviral transduction of OP9 cells (ATCC) with pMSCVpuro-hDLL4 construct (Clontech). For 3-dimensional (3D) culture, CD34+ cells were purified from mononuclear cells (purified by Ficoll) from either fresh or frozen and thawed umbilical cord blood (CB) or PB using the MACS kit (Miltenyi). CD34+ cells were mixed with trypsin-harvested OP9-DLL4 cells (1:23 mix) to form a 2.5-µL cell pellet, which was placed on a dry 25-mm polycarbonate culture insert (0.4 µm porosity, Nunc, catalog number 137060). This insert was transferred in a 6-well dish containing 1.5 mL media. Media was changed every 3 or 4 days. Cells were harvested from the insert for fluorescence-activated cell sorter analysis by vigorous pipetting. Detailed methods and reagents are provided in supplemental Methods and supplemental Table 2, respectively.
Results and discussion
We initially assessed the classical 2D-liquid culture system3 using limiting quantities of CB-derived and PB-derived CD34+ hematopoietic stem cells (HSCs). In our experience, we normally plate at least 5000 CD34+ cells per cm2 of culture surface area in order to successfully differentiate CD34+ cells into T cells in 2D-liquid culture. T-cell differentiation from PB-CD34+ HSCs was severely compromised when initial plating numbers were slightly decreased compared with equivalent numbers of CB-CD34+ HSCs (supplemental Figure 1A). Therefore, coculture of PB-CD34+ HSCs with OP9-DLL4 in monolayer is impractical given the small blood volume recommended in newborns or infants. We next placed a mixture of CD34+ HSCs and OP9-DLL4 cells onto a porous (0.4 µm) culture insert in a 3D culture system,4 identified as the air-liquid interface. This type of system was shown to produce a better differentiation of CD34+ HSCs in 3D compared with monolayer cultures, possibly due to the requirement of intricate cell-cell interactions only achieved in the 3D system, which recapitulates more closely the thymic architecture.4,5 By introducing significant modifications to this 3D culture system, we used limiting numbers of PB- or CB-CD34+ HSCs and obtained a successful and robust differentiation (supplemental Figure 1B) with as few as 500 CD34+ HSC. The originality of our culture system is that it allows to apply efficiently a very small number of cells (within a small volume) onto the inserts without inducing spreading, which is key for the formation of a tightly packed organoid. Notable differences with the previously published artificial thymic organoid system5 include the use of GFP-free OP9-DLL4 cells instead of OP9/MS5-DLL1-GFP cells, as well as fetal bovine serum–containing media and nonwettable culture inserts.
We applied our in vitro 3D T-cell differentiation assay to verify whether we could discern intrinsic from extrinsic HSC differentiation defects using limiting quantities of PB from young SCID patients. We reasoned that HSCs from patients harboring mutations that negatively impact their inherent differentiation capacity into T cells (IL2RG/γc, RAG1/2, and ADA) should not be able to produce advanced T-cell precursors. Conversely, HSCs from patients only affected in their thymic function (TBX1 or DiGeorge syndrome) should be able to properly differentiate into CD3+ T cells. As a demonstration of an intrinsic defect of differentiation, we showed that PB-CD34+ HSCs from an IL2RG/γc patient could not differentiate into CD34−CD7+CD1a+ double-negative (DN) cells, CD4+CD8+ double-positive (DP) cells, or CD3+ cells after 3 weeks of culture (Figure 1A), although CD34+CD7+ pro-T cells were abundantly present (data not shown). On the other hand, PB-CD34+ HSCs from a SCID patient with complete RAG2 deficiency (null mutation) advanced normally to the CD34−CD7+CD1a+ DN stage, with scarce presence of CD4+CD8+ DP cells (0.72% vs 22.7% for control) and no CD3+ cells after 5 weeks of culture (Figure 1B). An additional RAG1 hypomorph patient presenting with Omenn syndrome could however differentiate efficiently up to the CD4+CD8+ DP stage (39.5% vs 12.36% for control) after 5 weeks of culture but did not show evidence of CD3+ cell presence (Figure 1C).
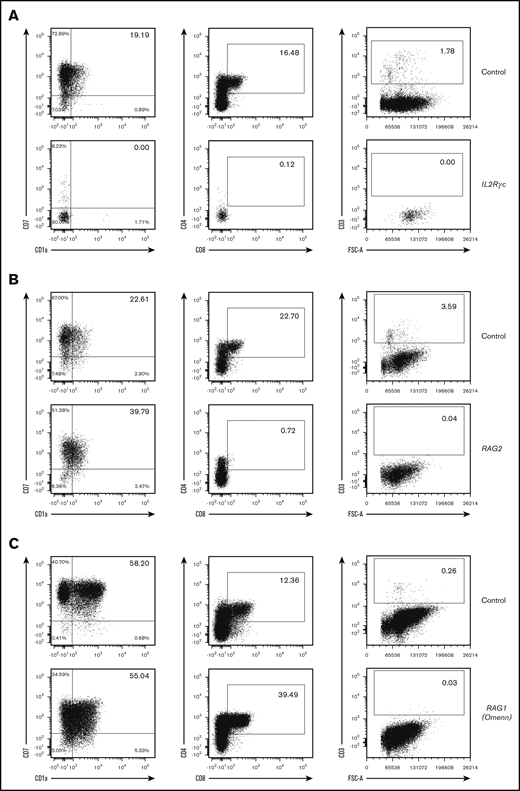
Abnormal in vitro T-cell differentiation of PB-CD34+cells with intrinsic deficiency. (A) A total of 1500 CD34+ cells from PB of an IL2RG/γc SCID patient and a control (cord blood) were differentiated in 3D culture for 3 weeks. (B) A total of 500 CD34+ cells from PB of a RAG (complete deficiency) SCID patient and control (cord blood) were differentiated in 3D culture for 5 weeks. (C) A total of 600 CD34+ cells from PB of a RAG (partial deficiency; Omenn syndrome) SCID patient and control (PB of healthy adult donor) were differentiated in 3D culture for 5 weeks. Displayed events are gated from single cells, negative for 7-AAD and positive for human CD45. Percentages among CD45+7-AAD−CD34−CD19− cells (left column) and CD45+7-AAD− cells (2 right columns) are indicated in inlets. FSC-A, forward scatter area.
Abnormal in vitro T-cell differentiation of PB-CD34+cells with intrinsic deficiency. (A) A total of 1500 CD34+ cells from PB of an IL2RG/γc SCID patient and a control (cord blood) were differentiated in 3D culture for 3 weeks. (B) A total of 500 CD34+ cells from PB of a RAG (complete deficiency) SCID patient and control (cord blood) were differentiated in 3D culture for 5 weeks. (C) A total of 600 CD34+ cells from PB of a RAG (partial deficiency; Omenn syndrome) SCID patient and control (PB of healthy adult donor) were differentiated in 3D culture for 5 weeks. Displayed events are gated from single cells, negative for 7-AAD and positive for human CD45. Percentages among CD45+7-AAD−CD34−CD19− cells (left column) and CD45+7-AAD− cells (2 right columns) are indicated in inlets. FSC-A, forward scatter area.
To assess the differentiation outcome of HSCs with extrinsic defects, we cultured PB-CD34+ HSCs from a patient with TBX1 mutation. As expected, HSCs differentiated normally up to the CD3+ cell stage, with the major progenitor intermediates (pro-T, DN, and DP) being present at levels similar to controls (Figure 2A), correctly identifying a problem outside of the HSC compartment, such as the thymus. The physicians caring for a patient with DiGeorge syndrome who had not developed T cells 1 year after thymus transplantation sent blood to determine if the patient also had a SCID mutation. The results (Figure 2B) indicated that the patient had a thymic defect, and subsequently, without receiving a HSC transplant (HSCT), recipient T cells did appear.
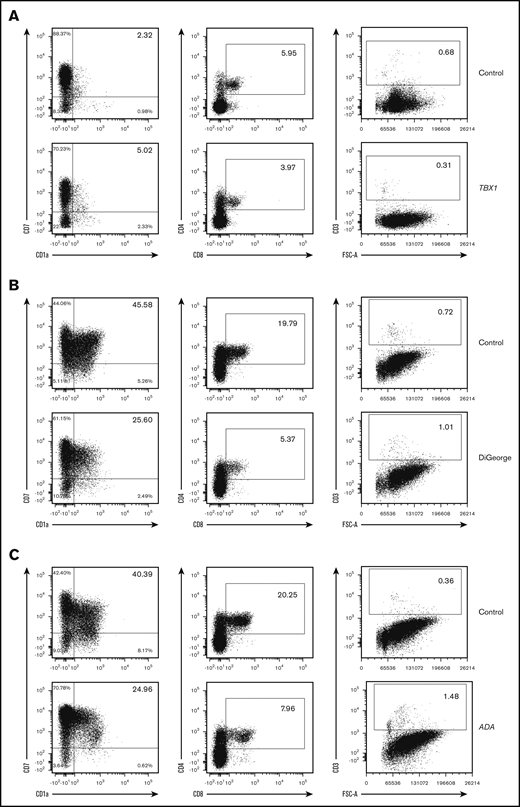
Normal in vitro T-cell differentiation of PB-CD34+cells from SCID patients with thymic defects and ADA deficiency. (A) A total of 5000 CD34+ cells from PB of a TBX1 SCID patient and control (cord blood) were differentiated in 3D culture for 5 weeks. (B) A total of 1900 CD34+ cells from PB of a DiGeorge syndrome SCID patient and control (PB of a healthy adult donor) were differentiated in 3D culture for 5 weeks. (C) A total of 2000 CD34+ cells from PB of an ADA SCID patient and control (PB of a healthy adult donor) were differentiated in 3D culture for 3 weeks. Displayed events are gated from single cells, negative for 7-AAD and positive for human CD45. Percentages among CD45+7-AAD−CD34−CD19− cells (left column) and CD45+7-AAD− cells (2 right columns) are indicated in inlets.
Normal in vitro T-cell differentiation of PB-CD34+cells from SCID patients with thymic defects and ADA deficiency. (A) A total of 5000 CD34+ cells from PB of a TBX1 SCID patient and control (cord blood) were differentiated in 3D culture for 5 weeks. (B) A total of 1900 CD34+ cells from PB of a DiGeorge syndrome SCID patient and control (PB of a healthy adult donor) were differentiated in 3D culture for 5 weeks. (C) A total of 2000 CD34+ cells from PB of an ADA SCID patient and control (PB of a healthy adult donor) were differentiated in 3D culture for 3 weeks. Displayed events are gated from single cells, negative for 7-AAD and positive for human CD45. Percentages among CD45+7-AAD−CD34−CD19− cells (left column) and CD45+7-AAD− cells (2 right columns) are indicated in inlets.
Surprisingly, PB-CD34+ HSCs of an adenosine deaminase (ADA)–deficient SCID patient differentiated normally up to the CD3+ cell stage (Figure 2C). The presence of OP9-DLL4 feeder cells and/or serum included in the culture media could have compensated in trans for the ADA deficiency. Another hypothesis is that the environment of the OP9-DLL4 assay results in lower levels of deoxyadenosine and toxic derivatives than in vivo in the thymus of these patients.6,7 This limitation of our test for ADA-deficient patients is balanced by the availability of specific metabolic/enzyme assessments that can easily make the diagnosis of ADA deficiency.
Our data are very similar to those of the companion paper from Bosticardo et al,8 where patients’ bone marrow (BM) samples were used. The DiGeorge and the RAG2-null BM samples were obtained from the same patients as for our study. However, for this common SCID patient with a RAG2-null mutation, Bosticardo et al did not observe a marked absence of DP cells, as was the case in our 5-week RAG-null PB-HSC cultures. Our data are, however, reconcilable if one considers the duration of the culture. Indeed, Brauer et al showed that induced pluripotent stem cell–derived CD34+ cells from patients with RAG mutations could differentiate early (3 weeks) into DP cells, but DP cells were then lost later on in the culture (5 weeks).9 In the accompanying article, Bosticardo et al report a major decrease of DP cells when the culture was prolonged to 6 weeks. This highlights that future assessment of SCID differentiation should take into account the dynamic nature of the results.
The cases described above are SCID with defined diagnosis. We also tested a “real” case in which the cause of SCID was undefined. This 2-year-old male had an urgent positive newborn screen for SCID and was followed for a chronic T-cell lymphopenia with low naive CD4+ cell counts but normal B and natural killer cell counts. He also had short stature and transfusion-dependent chronic anemia, with a paucity of erythroid progenitors in a BM aspirate. All diagnostic tests for both T-cell lymphopenia and the anemia were inconclusive, including DNA sequencing of a SCID gene panel, primary immunodeficiency panel, and whole exome. The differential diagnosis included an intrinsic HSC defect vs a thymic disorder. From a 5-mL PB sample, 6 million PB mononuclear cells were isolated, from which 436 CD34+ cells were purified and cocultured with OP9-DLL4 cells, as described above. As shown in supplemental Figure 2, after 5 weeks in T-cell differentiation conditions, very few CD4+CD8+ DP cells were observed, strongly suggesting an intrinsic defect that should require HSCT. A similar failure of in vitro T-cell generation occurred when CD34+ cells obtained from a BM sample were assayed in the Notarangelo laboratory (see companion article by Bosticardo et al8 ; data not shown). With these results, the patient received an HSCT from an 11/12 HLA-compatible unrelated donor. At 10 weeks after transplant, the patient demonstrated 100% donor chimerism with early development of donor T cells.
Overall, we have presented a proof-of-principle for an assay using cells obtained from a minimal volume of PB to inform the physician about the approximate level of deficiency (HSPC vs thymus defect) in unclassified SCID. The similarity of our data with those obtained from BM samples (see accompanying article by Bosticardo et al) adds to the robustness of our findings and demonstrates that assessing peripheral CD34+ cells in our 3D culture system is very similar to assessing BM-derived CD34+ cells. These data suggest that our T-cell differentiation assay, with as little as 5 mL PB, is robust and has a rapid readout (5 weeks). To address some of its shortcomings, further modifications of the assay would be to make it feeder-free (reduction in variability, avoidance of ADA secretion), as well as to combine it with ex vivo HSC expansion for repeated testing and genetic correction, ultimately helping us to seamlessly characterize novel SCID mutations.
Send data sharing requests via e-mail to the corresponding author, Elie Haddad (elie.haddad@umontreal.ca).
Acknowledgments
The authors thank members of the L. Notarangelo laboratory for helpful discussions and sharing results, including patient samples.
This work was supported by the Canadian Institutes of Health Research (grant 231438), The Bank of Montreal research chair in pediatric immunology, and by the Division of Allergy, Immunology and Transplantation, National Institute of Allergy and Infectious Diseases and the Office of Rare Diseases Research, National Center for Advancing Translational Sciences, National Institutes of Health, cooperative agreement U54-AI082973.
Authorship
Contribution: P.B. and E.H. designed the experiments and analyzed the data; P.B. and A.B.L.C. performed the experiments; P.B. and E.H. wrote the manuscript; S.-Y.P., M.L.M., N.H., J.M.P., J.A.C., J.W.L., and B.R.O. provided patient samples and comments on the manuscript; and all authors reviewed and approved the manuscript.
Conflict-of-interest disclosure: Cultured thymus tissue implantation (RVT-802) is an investigational product implanted into patients under an Investigational New Drug application with the US Food and Drug Administration. M.L.M. is the “sponsor” of the investigations and developed the technology for cultured thymus tissue implantation. Duke has licensed the technology to Enzyvant Therapeutics. M.L.M. and Duke have received royalties from Enzyvant. Portions of M.L.M.’s and her research team’s salaries are being paid by funding from Enzyvant. If the technology is commercially successful in the future, M.L.M. and Duke may benefit financially. The remaining authors declare no competing financial interests.
Correspondence: Elie Haddad, Department of Pediatrics, University of Montreal, Immunology Division, CHU Sainte-Justine, 3175 Cote Sainte-Catherine, Room 6.17.003, Montreal, QC H3T 1C5, Canada; e-mail: elie.haddad@umontreal.ca.

