

In this issue of Blood, Feitsma et al report the structure of platelet glycoprotein VI (GPVI) complexed with collagen-related peptides and propose the structural basis of GPVI clustering on collagen fibers.1
GPVI has raised interest as a potential target for effective and safe antithrombotic molecules. GPVI belongs to the immunoglobulin (Ig) receptor superfamily that includes killing inhibitory receptors, leukocyte Ig–like receptors, FcαR1, and leukocyte-associated Ig-like receptors (LAIRs). GPVI is a tyrosine-based activation motif (ITAM) immunoreceptor because of its association with the Fc-receptor γ-chain (FcRγ).
The ectodomain of GPVI consist of 2 Ig-like domains (D1 and D2) and a glycosylated stalk region. GPVI is embedded in the membrane through a single transmembrane helix bridged to the FcRγ chain, and it has a short intracellular tail. Binding of the extracellular region to collagen results in intracellular signaling and platelet activation. GPVI has been crystalized in combination with different ligands by several different groups. All structural studies have focused on the D1 and D2 domains,2,3 but the structure of GPVI complexed with collagen has not previously been reported.
In their article, Feitsma et al are the first to report the structure of D1 and D2 in complex with triple helical collagen-related peptides (CRPs) containing 3 or 5 stretches of the GPVI binding motif Gly-Pro-hydroxyPro. The ligand-bound structures reveal binding through the D1 domain. Unexpectedly, the CRP binding site is some distance away from the previously modeled collagen binding site,2 but it is consistent with the previously reported collagen binding site on LAIR-1.4 These findings demonstrate the methodologic advantage of crystallography, which has less risk of error than in silico docking. CRP binding does not result in significant structural changes to the GPVI structure and suggests that collagen binding does not induce a conformational change resulting in an active GPVI state.
A collagen fiber is formed by individual triple helices that assemble to form a complex fibrous structure. Platelet activation by collagen requires clustering of multiple copies of GPVI-FcRγ complexes that lead to the phosphorylation of the FcRγ chain and downstream signaling. Feitsma et al use their structure to propose models for how multiple GPVI subunits can cluster along a parallel fiber at the level of the GPVI-specific binding motifs (see figure). They demonstrate that a large separation between GPVI subunits is required (32-40 Å). In addition, looking at simultaneous binding to parallel helices, the rigid angle between D1 and D2 imposes steric constraints, which means that it is not feasible for a second GPVI subunit to bind parallel helices less than 34 Å apart. This allows us to make basic predictions regarding how GPVI clustering along a collagen fiber might appear. As an extreme example, a platelet with a diameter of 3 μm would allow for the clustering of close to 900 GPVI receptors from this one platelet along one fiber. This is a simplified model because it does not consider the presence of FcRγ, the distribution of GPVI in the membrane, or the spacing between GPVI binding sites along the fiber, but we can begin to visualize what this might look like on the platelet surface.
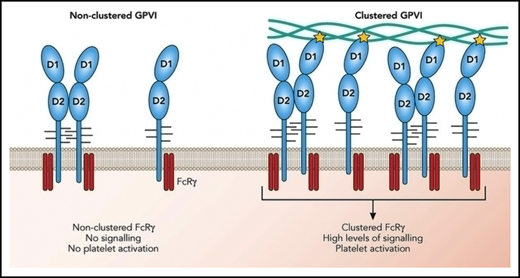
GPVI activation requires receptor clustering but not dimerization. On resting platelets, GPVI monomers (shown in blue) are present on the platelet surface co-expressed with the FcRγ chain (red). A limited proportion of GPVI is present as dimers. GPVI clustering along a collagen fiber increases the local concentration of the FcRγ chain resulting in amplified signaling events leading to platelet activation. The role of dimerization is to further increase the concentration of FcRγ. The yellow star represents an interaction with collagen and highlights that only 1 GPVI within a dimer can interact with collagen. Professional illustration by Patrick Lane, ScEYEnce Studios.
GPVI activation requires receptor clustering but not dimerization. On resting platelets, GPVI monomers (shown in blue) are present on the platelet surface co-expressed with the FcRγ chain (red). A limited proportion of GPVI is present as dimers. GPVI clustering along a collagen fiber increases the local concentration of the FcRγ chain resulting in amplified signaling events leading to platelet activation. The role of dimerization is to further increase the concentration of FcRγ. The yellow star represents an interaction with collagen and highlights that only 1 GPVI within a dimer can interact with collagen. Professional illustration by Patrick Lane, ScEYEnce Studios.
The findings by Feitsma et al also shed light on the still debated question of GPVI dimerization vs clustering. Binding of antibodies that have a higher affinity for GPVI dimers than they have for monomers has been interpreted as reflecting the presence of platelet surface-expressed dimeric GPVI.5,6 Platelets labeled with these antibodies increase in patients with cardiovascular risk factors,5,7 suggesting that GPVI dimerization primes platelets. GPVI dimerization has also been observed on transfected cells.8 Consistent with these findings, all crystal structures of GPVI have revealed dimeric conformations of GPVI, and the structures described by Feitsma et al reveal the same back-to-back dimer conformation reported by Horii et al2 with sites of dimerization some distance away from the collagen binding site.
Combining all the known dimer conformations, such GPVI dimers are unable to support simultaneous binding to 2 parallel collagen fibers. Therefore, the dimeric GPVI structures do not support the idea that collagen binding requires a dimeric conformation, unless GPVI dimerization on the cell surface adopts a conformation different from those in the crystal structures. These results support the proposition that GPVI clustering is the important event for triggering downstream signaling, as shown by the fact that soluble collagen is unable to activate platelets. Conversely, these data raise the question of an inside-out signaling that would be able to control the avidity of GPVI for collagen, as has been observed more generally for other FcRs.9 For now, the exact process for the formation of GPVI dimers remains unknown, but it could involve orientating the FcRγ chains into signal active states or simply could bring more FcRγ chains to the cluster. Feitsma et al also positioned binding motifs of von Willibrand factor (VWF) and integrin α2β1 on fibrillar collagen III. Interestingly, the frequent distribution of GPVI binding motifs and the relatively sparse VWF and α2β1 binding sites could support the simultaneous binding of GPIb, integrin α2β1, and GPVI consistent with the known cross-talk between these receptors.10
Now that the binding site has been mapped, we are better able to design and screen for selective inhibitors that directly inhibit collagen binding. The limitation of such molecules could be that they will preserve GPVI interaction with its other ligands such as fibrin and thus have a limited efficiency in thrombus growth. With the increased knowledge of GPVI clustering mechanisms, we can also design inhibitors that do not bind close to the collagen binding site but still sterically occlude the collagen fiber downstream.
More studies are now required to determine the structure of full-length GPVI anchored in the membrane in complex with the FcRγ chain. This will allow us to determine the GPVI conformation in its biological setting and compare that structure to the crystal structures. It will also allow us to map the interactions between GPVI and FcRγ and how binding to extracellular GPVI results in intracellular signaling through FcRγ. Techniques such as cryoelectron microscopy may provide valuable tools for studying this.
Conflict-of-interest disclosure: M.J.-P. is a founder and scientific adviser for Acticor Biotech. A.S. declares no competing financial interests.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal