

Key Points
The proteomic landscape of mutant GATA1, GFI1B, and RUNX1 platelets identified more than 300 differentially expressed proteins compared with controls.
GATA1, GFI1B, and RUNX1 mutant platelets have reduced expression of numerous coagulation proteins, but none of these are common to all transcription factor mutant platelets.

Abstract
Mutations in the transcription factors GATA binding factor 1 (GATA1), growth factor independence 1B (GFI1B), and Runt-related transcription factor 1 (RUNX1) cause familial platelet and bleeding disorders. Mutant platelets exhibit common abnormalities including an α-granule reduction resulting in a grayish appearance in blood smears. This suggests that similar pathways are deregulated by different transcription factor mutations. To identify common factors, full platelet proteomes from 11 individuals with mutant GATA1R216Q, GFI1BQ287*, RUNX1Q154Rfs, or RUNX1TD2-6 and 28 healthy controls were examined by label-free quantitative mass spectrometry. In total, 2875 platelet proteins were reliably quantified. Clustering analysis of more than 300 differentially expressed proteins revealed profound differences between cases and controls. Among cases, 44 of 143 significantly downregulated proteins were assigned to platelet function, hemostasis, and granule biology, in line with platelet dysfunction and bleedings. Remarkably, none of these proteins were significantly diminished in all affected cases. Similarly, no proteins were commonly overrepresented in all affected cases compared with controls. These data indicate that the studied transcription factor mutations alter platelet proteomes in distinct largely nonoverlapping manners. This work provides the quantitative landscape of proteins that affect platelet function when deregulated by mutated transcription factors in inherited bleeding disorders.
Introduction
Mutations in the transcription factors (TFs) GATA binding factor 1 (GATA1), growth factor independence 1B (GFI1B), and Runt-related transcription factor 1 (RUNX1) cause inherited platelet and bleeding disorders with highly overlapping megakaryocyte and platelet phenotypes.1-5 The bleeding diathesis is caused by thrombocytopenia associated with dysfunctional platelets that exhibit a shortage in α-granules. The phenotypic similarities suggest that the TFs regulate common gene programs. We used label-free quantitative mass spectrometry to reveal downstream effects of autosomal dominant GATA1R216Q, GFI1BQ287*, RUNX1Q154Rfs, and RUNX1TD2-6 mutations on platelets from affected cases vs healthy controls. These mutations encompass DNA binding domains. GATA1R216Q might result in a gain of function, based on the analogous GATA2R307W mutant that predisposes to myeloid leukemia.6 GFI1BQ287* functions in a dominant-negative manner, whereas the effect of the studied RUNX1 mutations (frameshift and tandem duplication) are unknown.2,3 Here, we identified significantly enriched and depleted proteins in TF mutant platelets and determined whether these converge to specific biological pathways.
Study design
Platelets were isolated from blood from individuals with different gray-platelet–associated TF mutations: GATA1R216Q (n = 2), GFI1BQ287* (n = 6), RUNX1Q154Rfs (n = 1), and RUNX1TD2-6 (n = 2) (tandem duplication exon 2-6) and healthy controls (n = 28) (supplemental Table 1, available on the Blood Web site). Platelet lysates were prepared for mass spectrometry as described and measured in triplicate.7 This study was approved by the Medical Ethics Committee of the Radboud UMC and conducted in accordance with the Declaration of Helsinki. Platelet proteomes were subjected to mass spectrometry (Orbitrap Fusion Tribrid mass spectrometer, Nanospray Flex Ion Source, Thermo Fisher Scientific) and analyzed using Xcalibur and Maxquant software (version 1.5.2.8). See supplemental Data for details.
Results and discussion
Mutant TFs significantly affect platelet proteomes
A total of 2875 primary platelet proteins were reliably quantified, of which 313 (11%) were significantly differentially expressed between mutant cases vs controls (supplemental Table 2A-E). Among affected cases, GATA1 and RUNX1 mutant and control cases were more similar with each other compared with GFI1B cases (Figure 1A). K-means clustering analysis separated healthy controls from mutant proteomes, whereas individuals with the same mutation clustered together except for 1 GFI1BQ287* case (Figure 1B). Cluster 1 was predominantly characterized by a reduction in protein levels in ≥1 of the cases compared with controls. Clusters 2 and 3 contained upregulated proteins in, respectively, the RUNX1 and GATA1 mutants and the GFI1BQ287* vs controls. To zoom in on proteome differences, we performed cluster analyses per mutant TF compared with controls, confirming the marked changes for GFI1BQ287* compared with other mutants (supplemental Figure 1; supplemental Table 2B-E).
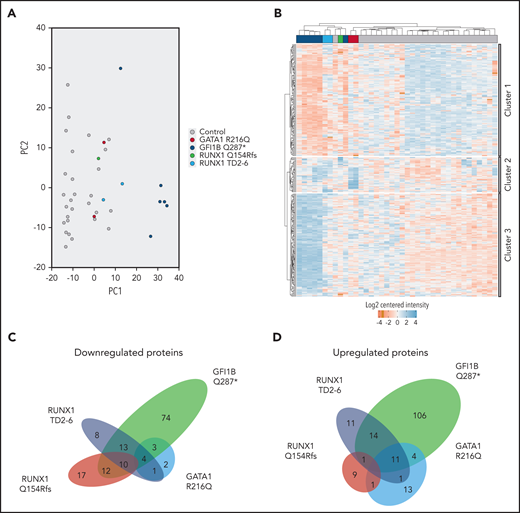
Differentially expressed proteins make up the transcription factor mutant platelet proteome. (A) Principal component analysis shows clustering of control samples from 28 individuals, clearly separated from GFI1BQ287* mutant-derived platelets (PC1). RUNX1 and GATA1 mutant platelets are more similar to each other and controls compared with GFI1B mutant platelets. (B) Hierarchical clustering was observed between control and mutant-derived platelets by K-means clustering analysis. One GFI1B case maps between a GATA1 and RUNX1 case and 1 control case maps between 2 RUNX1 cases. The differentially expressed proteins were subdivided into 3 clusters based on their expression similarity. Cluster 1 represents proteins that are mainly downregulated in mutants compared with controls. Cluster 2 represents proteins that are upregulated in GATA1R216Q and RUNX1 mutants compared with controls. Cluster 3 shows upregulated proteins for GFI1BQ287* mutant platelets. Downregulated proteins are marked in red; upregulated proteins are marked in blue. (C) Venn diagram from downregulated proteins shows overlap on affected proteins between TF mutants, but none are common to all TFs. Numbers in the different areas indicate the amount of shared proteins between cases. Proteins converging to significantly underrepresented processes are listed in supplemental Tables 2F and 4. (D) Venn diagram from upregulated proteins shows overlap of common upregulated proteins, but none are common to all TFs. Proteins converging to significantly overrepresented processes are listed in supplemental Tables 2H and 4. The total number of proteins down- and upregulated for RUNX1 variants was 65 + 47 = 112, whereas only 10 + 1 = 11 proteins were in common.
Differentially expressed proteins make up the transcription factor mutant platelet proteome. (A) Principal component analysis shows clustering of control samples from 28 individuals, clearly separated from GFI1BQ287* mutant-derived platelets (PC1). RUNX1 and GATA1 mutant platelets are more similar to each other and controls compared with GFI1B mutant platelets. (B) Hierarchical clustering was observed between control and mutant-derived platelets by K-means clustering analysis. One GFI1B case maps between a GATA1 and RUNX1 case and 1 control case maps between 2 RUNX1 cases. The differentially expressed proteins were subdivided into 3 clusters based on their expression similarity. Cluster 1 represents proteins that are mainly downregulated in mutants compared with controls. Cluster 2 represents proteins that are upregulated in GATA1R216Q and RUNX1 mutants compared with controls. Cluster 3 shows upregulated proteins for GFI1BQ287* mutant platelets. Downregulated proteins are marked in red; upregulated proteins are marked in blue. (C) Venn diagram from downregulated proteins shows overlap on affected proteins between TF mutants, but none are common to all TFs. Numbers in the different areas indicate the amount of shared proteins between cases. Proteins converging to significantly underrepresented processes are listed in supplemental Tables 2F and 4. (D) Venn diagram from upregulated proteins shows overlap of common upregulated proteins, but none are common to all TFs. Proteins converging to significantly overrepresented processes are listed in supplemental Tables 2H and 4. The total number of proteins down- and upregulated for RUNX1 variants was 65 + 47 = 112, whereas only 10 + 1 = 11 proteins were in common.
Many proteins implicated in platelet homeostasis are underrepresented in TF mutant platelets
Because platelet dysfunction is related to α-granule shortage in cases with TF mutations, we first focused on 144 proteins significantly less abundant in 1 or more TF variants compared with controls. The most significantly affected biological pathways represented by these proteins were involved in hemostasis, platelet degranulation, response to cytosolic Ca2+, and platelet activation, signaling, and aggregation (supplemental Table 2F). Most of these proteins were present in cluster 1 and contained well-known α-granule proteins such as von Willebrand factor, thrombospondin, and platelet factor 4 (supplemental Tables 2F and 3A-B). Apart from known coagulation proteins, 100 proteins previously unknown to be diminished in these bleeding disorders were identified (supplemental Table 2F). These proteins associated with pathways like extracellular matrix organization, cell surface interactions at the vascular wall, and laminin interactions. It will be important to determine how the reduced expression of the 144 proteins contribute to bleeding. Finally, we analyzed proteins that were not classified as being significantly differentially expressed between cases and controls. This identified more than 150 hemostasis proteins, including GP9, PDGFA, and ITGA2B (supplemental Table 2A-E,G).
TF mutation-specific underrepresentation of biological pathways
Despite a broad underrepresentation of proteins involved in platelet aggregation for each individual TF mutation, none of these were affected in all the studied cases. In fact, a large number of proteins (n = 101) with significant lower abundance was unique to any of the TF variants (Figure 1C). In GFI1BQ287* platelets, 74 proteins were uniquely underrepresented, whereas for the RUNX1Q154fs, RUNX1TD2-6, and GATA1R216Q mutants, respectively 17, 8, and 2 proteins were uniquely downregulated. In total, 43 proteins were shared between 2 or more TF mutant cases, of which 10 were shared among GFI1BQ287*, RUNX1Q154Rfs, and RUNX1TD2-6 cases compared with controls. MYL9, present among this latter group, has been proposed to be directly regulated by RUNX1 (supplemental Tables 2A-E and 4).8 Remarkably, of 112 differentially expressed RUNX1 mutant proteins, only 11 were shared between the 2 RUNX1 variants (Figure 1C-D). Apparently, the RUNX1 frameshift vs duplication mutation affects proteomes in different manners.
Differential representation of granule proteins in GATA1-, GFI1B-, and RUNX1-mutated platelets
A reduction of platelet α-granules is a hallmark of the studied bleeding disorders. Earlier mass spectrometry studies identified more than 800 platelet granule-associated proteins.9,10 Approximately half the amount of these granule proteins were detected and not differentially expressed between cases and controls. Compared with controls, 13 and 48 proteins were overrepresented and downmodulated, respectively (supplemental Table 5; Figure 2A). For the latter category, both α-granule content and membrane proteins were observed. These analyses pinpoint the paucity of α-granule proteins specific for the different TF mutants. We also determined whether any of the differentially expressed proteins are implicated in inherited bleeding disorders.11-13 This identified 7 underrepresented proteins, including F5 and F13A1 (Figure 2B; supplemental Table 6). As mutations in these proteins impair their function, their underrepresentation might contribute to platelet dysfunction in the studied bleeding disorders.
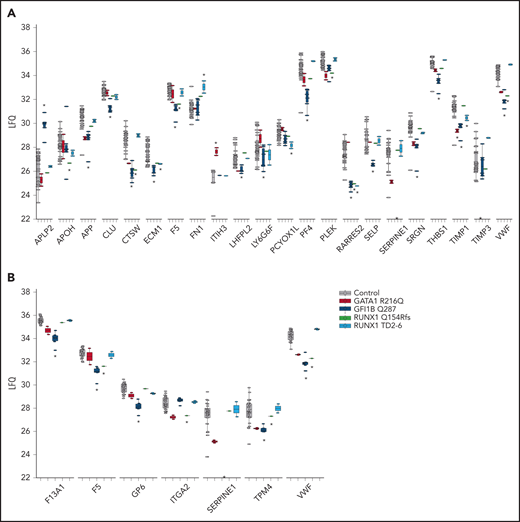
Significantly affected granule and bleeding disorder proteins. (A) Raw label free quantitation (LFQ) intensities were plotted for Reactome defined granule proteins for controls and each TF mutant. See supplemental Table 5 for complete list of granule associated proteins that were identified in platelet granule proteomic studies.9,10 Asterisks above or below the plotted LFQ values represent significantly differential expressed proteins to be up- and downregulated, respectively. The asterisks on the x-axis indicates complete absence of SERPINE1 in GFI1B mutant cases and TIMP3 in GATA1 mutant cases. (B) Raw LFQ intensities were plotted for inherited bleeding disorder associated proteins for controls and each TF mutant. Asterisks represent significantly downregulated proteins. The asterisk on the x-axis indicates complete absence of SERPINE1 in GFI1B mutant cases.
Significantly affected granule and bleeding disorder proteins. (A) Raw label free quantitation (LFQ) intensities were plotted for Reactome defined granule proteins for controls and each TF mutant. See supplemental Table 5 for complete list of granule associated proteins that were identified in platelet granule proteomic studies.9,10 Asterisks above or below the plotted LFQ values represent significantly differential expressed proteins to be up- and downregulated, respectively. The asterisks on the x-axis indicates complete absence of SERPINE1 in GFI1B mutant cases and TIMP3 in GATA1 mutant cases. (B) Raw LFQ intensities were plotted for inherited bleeding disorder associated proteins for controls and each TF mutant. Asterisks represent significantly downregulated proteins. The asterisk on the x-axis indicates complete absence of SERPINE1 in GFI1B mutant cases.
TF mutation-specific overrepresentation of biological pathways
Despite a shortage of α-granules proteins, clustering analyses identified 170 proteins to be significantly upregulated in mutant platelets vs controls. Similar to downmodulated proteins, most of these proteins (n = 139) were specific for each of the TF mutants, and none of the proteins were common to all TF mutant platelets (Figure 1D). A total of 83 overrepresented biological pathways were identified, with erythrocyte-associated pathways, metabolism, and vascular endothelial growth factor signaling being most significant (supplemental Table 2H). Overrepresented proteins specific for GATA1R216Q, GFI1BQ287*, RUNX1Q154Rfs, and RUNX1TD2-6 variants were implicated in metabolism, cell communication, signaling, and matrix organization, respectively (supplemental Table 4). Eleven proteins were shared among GATA1R216Q, GFI1BQ287*, and RUNX1TD2-6 cases compared with controls, of which 5 proteins converge to metabolism (supplemental Tables 2A-E and 4).
The data presented here show that GATA1, GFI1B, and RUNX1 TF mutations affect platelet proteomes in largely distinct nonoverlapping manners. This could be caused by differential GATA1, GFI1B, and RUNX1 gene expression during megakaryocyte development.14,15 Alternatively, the observed proteomic differences might occur through differential binding of GATA1, GFI1B, and RUNX1 to the respective genes. However, we did not find specific differences of GATA1, GFI1B, and RUNX1 binding motifs in the promoters of genes encoding the 106 uniquely overrepresented and 74 underrepresented proteins in GFI1BQ287* platelets nor in the genes of over- or underrepresented proteins in any of the TF mutants (supplemental Figure 2). GATA1, GFI1B, and RUNX1 DNA binding in megakaryo-erythroblast K562 cells and primary megakaryocytes did not explain proteomic differences either (supplemental Figure 3).16-18
Together, this work provides quantitative insights in the proteomic abnormalities downstream of mutated TFs in developing megakaryocytes, which is reflected in the platelets they produce. It represents a framework for further studies in platelet biology. Although megakaryocyte and platelet abnormalities in these bleeding disorders are reminiscent, the proteome composition is different, indicating different roles for the TFs in platelet biology.
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD020343.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
This work was supported by the Landsteiner Foundation for Blood Transfusion Research (project 1531). A.E.M. is sponsored by US Department of Defense Horizon Award W81XWH2010904.
Authorship
Contribution: M.G.J.M.V.B., A.B.M., and B.A.V.d.R. designed and coordinated research; M.G.J.M.V.B., A.E.M., A.J.H., J.H.A.M., and F.P.J.V.A. collected and analyzed data; B.A.P.L.-V.G., J.J.W.M.J., and M.H. arranged patients and informed consent; E.V.d.A., A.S., S.A.D.M., and J.H.J. evaluated results; and M.G.J.M.V.B. and B.A.V.d.R. wrote the manuscript that was critically revised by all authors.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: A. B. Meijer, Plesmanlaan 125, 1066 CX, Amsterdam, The Netherlands; e-mail: s.meijer@sanquin.nl; and B. A. Van der Reijden, Geert Grooteplein zuid 10, 6525 GA Nijmegen, The Netherlands; e-mail: bert.vanderreijden@radboudumc.nl.
