

In this issue of Blood, show an unexpected role for biogenesis of lysosomal related organelle 2 (BLOC-2) in the formation of Weibel-Palade bodies (WPBs), a crucial storage compartment for hemostatic and inflammatory cargo in endothelial cells.1 The BLOC-2 complex is composed of 3 distinct subunits, HPS3, HPS5, and HPS6, which are absent in patients suffering from corresponding subtypes of Hermansky-Pudlak syndrome (types 3, 5, and 6).2 Patients with mutations in 1 of the BLOC-2 subunits have a severe bleeding tendency, consistent with the aberrant generation of storage compartments in platelets and endothelial cells.3 Ultrastructural analysis of platelets from patients with BLOC-2 deficiency revealed that BLOC-2 is crucial for the generation of dense, but not α granules in platelets.4 Similarly, studies focusing on the endothelial storage compartment in “ruby eye” (HPS6-deficient) mice showed an altered tubular organization of von Willebrand factor (VWF), which resulted in short and round WPBs releasing smaller VWF filaments.5 The current study confirms the effect of HPS6 on WPB morphology and provides evidence for the crucial role of BLOC-2 in CD63 trafficking to WPBs. CD63 is a notorious “late arrival” during WPB maturation because it is only trafficked to WPBs once they have budded from the trans-Golgi network.6,7 CD63 transport to WPBs is dependent on the integrity of the AP-3 complex.7,8 Very elegant cryo-EM studies have shown that CD63 resides in membrane bound luminal vesicles within WPBs.9
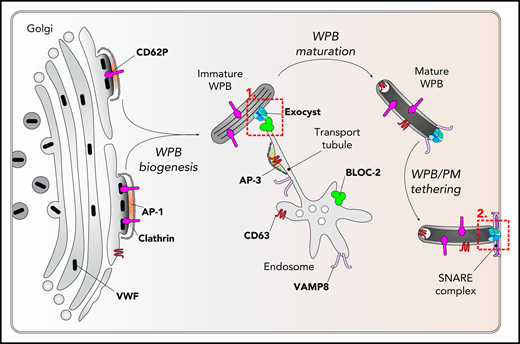
BLOC-2-mediated cargo delivery to maturing Weibel-Palade bodies. Immature Weibel-Palade bodies are generated in an AP-1/clathrin-dependent pathway by packaging of VWF quanta (black capsules), CD62P, and other cargo in the trans-Golgi network. Immature WPBs acquire CD63 in an AP-3-dependent pathway from endosomes. The findings of Sharda et al show that complex-formation between BLOC-2 and the exocyst is needed for transporting CD63 to immature WPBs (dashed red box 1). Fully mature WPBs are tethered to the plasma membrane through the exocyst complex (dashed red box 2). Sharda et al propose that the exocyst acts as a “clamp” preventing premature release of WPBs. Fusion of WPBs is mediated by so-called SNARE complexes. Recent findings have shown that SNARE proteins such as VAMP8 are recruited onto maturing WPBs through the same BLOC-2/AP-3-dependent pathway that is used by CD63.
BLOC-2-mediated cargo delivery to maturing Weibel-Palade bodies. Immature Weibel-Palade bodies are generated in an AP-1/clathrin-dependent pathway by packaging of VWF quanta (black capsules), CD62P, and other cargo in the trans-Golgi network. Immature WPBs acquire CD63 in an AP-3-dependent pathway from endosomes. The findings of Sharda et al show that complex-formation between BLOC-2 and the exocyst is needed for transporting CD63 to immature WPBs (dashed red box 1). Fully mature WPBs are tethered to the plasma membrane through the exocyst complex (dashed red box 2). Sharda et al propose that the exocyst acts as a “clamp” preventing premature release of WPBs. Fusion of WPBs is mediated by so-called SNARE complexes. Recent findings have shown that SNARE proteins such as VAMP8 are recruited onto maturing WPBs through the same BLOC-2/AP-3-dependent pathway that is used by CD63.
The current study of Sharda et al provides new insights into the HPS6-dependent trafficking of CD63 from endosomes to WPBs. Long endosomal tubular extensions terminating on WPBs were observed; remarkably, these endosomal tubules were misdirected in HPS6-deficient cells. These intriguing findings point toward a role of the BLOC-2 complex in tethering endosomal tubular extensions to WPBs. Using proteomic approaches, components of the exocyst complex, EXOC2 (sec5), EXOC4 (sec8), and EXO70, were identified as potential partners for the BLOC-2 complex. The exocyst is an octameric complex composed of 2 modules (vesicle-localized EXOC1-4 and target membrane-localized EXOC5-8), which are held together by several α-helical bundles, known as the “core of exocyst.”10 Knockdown of EXOC4 in endothelial cells phenocopied the HPS6 phenotype yielding immature, rounded WPBs that lack CD63. The findings strongly suggest that the BLOC-2 and exocyst complex both operate in the same pathway.
Apart from WPB maturation, the authors also addressed the potential role of BLOC-2 as well as the exocyst complex on agonist-induced release of VWF from WPBs. In line with previous studies, depletion of HPS6 both quantitatively and qualitatively impaired agonist-induced VWF release. The amount of VWF released in HPS6-depleted cells was reduced when compared with controls. Also, the proportion of high-molecular-weight VWF multimers was reduced in HPS6-depleted cells.
Unexpectedly, downregulation of EXOC4 enhanced agonist-induced secretion of VWF. Similar results were obtained using the EXO70 inhibitor Endosidin2, which also increased agonist-induced release of VWF. The molecular basis of these interesting, but somewhat counterintuitive, observations deserve further study.
Overall, the findings reported by Sharda et al highlight the crucial and novel role of the BLOC-2 complex in regulating the transport of cargo from endosomes to maturing WPBs.
The current study firmly implicates BLOC-2 and the exocyst complex in CD63 trafficking from endosomes to maturing WPBs. It is likely that CD63 delivery via this pathway is controlled by the AP3 complex. How AP3 and BLOC-2-dependent cargo delivery is coordinated remains to be established. The data contained in the current paper strongly suggest that BLOC-2/exocyst interactions are needed to direct endosomal tubular extensions to maturing WPBs, whereas AP-3 may control cargo selection and retention during formation and delivery of endosomal-derived transport tubules (see figure).
We have previously shown that VAMP8, a v-SNARE protein and part of the WPB exocytotic machinery, is transported to the WPB, “piggybacking” on the AP-3-dependent CD63 route.8 Mistrafficking of VAMP8 in AP-3-deficient endothelial cells derived from HPS2 patients results in reduced exocytotic competence,8 as also reported in this study, for endothelial cells that lack BLOC-2. Based on these data, it is likely that the BLOC-2/exocyst-dependent pathway described by Sharda et al is not only used for cargo delivery, but may also direct SNAREs, as well as tethering complexes to WPBs to successfully prime these organelles for rapid release of inflammatory and hemostatic cargo upon vascular triggering.
This stimulating study has significantly advanced our understanding of the formation of WPBs and paves the way for the identification of novel BLOC-2-regulated pathways that control delivery of cargo to storage compartments present in platelets and endothelial cells.
Conflict-of-interest disclosure: The authors declare no competing financial interests.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal