

Background: Proteolytic cleavage of von Willebrand factor (VWF) by a plasma metalloprotease ADAMTS13 is critical for normal hemostasis. Our previous studies demonstrate that binding of coagulation factor VIII (FVIII) through its light chain to VWF significantly accelerates the proteolytic cleavage of VWF by ADAMTS13 under fluidic shear. However, the mechanism underlying the FVIII-cofactor activity enhancing VWF proteolysis by ADAMTS13 under shear is not fully understood. Present study aims to test a hypothesis that FVIII binding to the D'D3 and/or other adjacent domains may facilitate a mechanic-induced conformational change in the central A2 domain of VWF, resulting in an increased accessibility and the cleavage of Tyr1605-Met1606 bond by ADAMTS13.
Methods: Recombinant B-domainless FVIII and various VWF fragments (D'D3, D'D3A1, D'D3A1A2, and A2) with an Avi-His-tag on their N-termini and a Spy-tag on their C-termini were expressed and affinity-purified to homogeneity. Laser optical tweezers analysis determined the force-induced conformational changes of VWF fragments in the absence or presence of FVIII. Atomic force microscopy (AFM) and surface plasmon resonance (SPR) determined the binding events and kinetics between FVIII and various VWF domains.
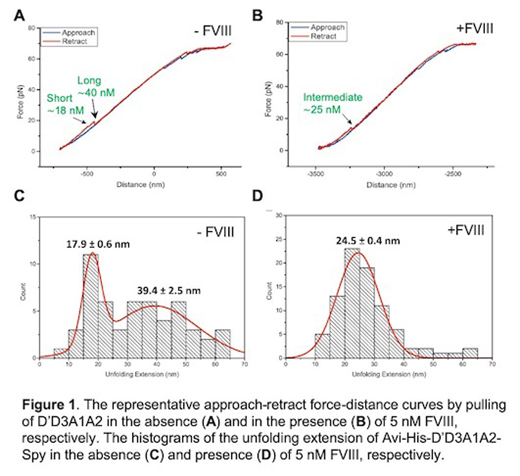
Results: Laser optical tweezers analysis demonstrated a Gaussian distribution of unfolding extension events upon pulling the single D'D3A1 fragment with the most probable extension distance of 22.8 ± 1.9 nm (mean ± SEM). Addition of 5 nM FVIII to the D'D3A1 or D'D3 fragment resulted in only a small decrease (~25%) or no change in the pulling extension profile. However, two peaks of Gaussian distribution were detected when pulling the D'D3A1A2 with one peak at ~18 nm and the other at ~40 nm. Addition of 5 nM FVIII to the D'D3A1A2 fragment, only one unfolding event was detected with the most probable unfolding extension distance of 24.5 ± 0.4 nm. Moreover, a single Gaussian distribution of unfolding extension profile was detected when pulling an isolated A2 fragment with the most probable unfolding extension of 31 ± 1 nm. Interestingly, addition of 5 nM FVIII resulted in a significant reduction of the most probable unfolding extension to 16.0 ± 0.4 nm, suggesting that FVIII may directly interact with A2 domain. AFM and SPR demonstrated the binding kinetics between FVIII and various VWF fragments. As shown, the intrinsic off-rate (k0) between FVIII and D'D3 was 0.19 ± 0.9 s-1 and the barrier width (Δx‡) between the naive state and the transition state was 1.5 ± 0.2 Å; the k0 between FVIII and D'D3A1A2 was 2.8 ± 1.0 s-1 and the barrier width (Δx‡) between the naive state and the transitional state was 1.7 ± 0.2 Å. Surprisingly, there was a detectable, but relatively weak binding between FVIII and A2 domain using AFM. SPR experiments confirmed the binding of FVIII to D'D3, D'D3A1A2, and A2 with the dissociation constant (KD) of ~2.9 nM, ~5.2 nM and ~1.6 nM, respectively. These results demonstrate that the specific interaction between FVIII and D'D3 or A2 domain.
Conclusions: We have demonstrated for the first time using single-molecule force spectroscopy that binding of FVIII to the D'D3 and/or A2 domain of VWF may facilitate the force-induced conformational changes of the A2 domain, resulting in the enhanced proteolysis by ADAMTS13 under force. Our findings shed new light on how FVIII may regulate VWF proteolysis and function under (patho) physiological conditions.
Zheng:Shire/Takeda: Research Funding; Alexion: Speakers Bureau; Clotsolution: Other: Co-Founder; Ablynx/Sanofi: Consultancy, Speakers Bureau.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal