

Abstract
The culminating step of the intraerythrocytic development of Plasmodium falciparum, the causative agent of malaria, is the spectacular release of multiple invasive merozoites on rupture of the infected erythrocyte membrane. This work reports for the first time that the whole process, taking place in time scales as short as 400 milliseconds, is the result of an elastic instability of the infected erythrocyte membrane. Using high-speed differential interference contrast (DIC) video microscopy and epifluorescence, we demonstrate that the release occurs in 3 main steps after osmotic swelling of the infected erythrocyte: a pore opens in ∼ 100 milliseconds, ejecting 1-2 merozoites, an outward curling of the erythrocyte membrane is then observed, ending with a fast eversion of the infected erythrocyte membrane, pushing the parasites forward. It is noteworthy that this last step shows slight differences when infected erythrocytes are adhering. We rationalize our observations by considering that during the parasite development, the infected erythrocyte membrane acquires a spontaneous curvature and we present a subsequent model describing the dynamics of the curling rim. Our results show that sequential erythrocyte membrane curling and eversion is necessary for the parasite efficient angular dispersion and might be biologically essential for fast and numerous invasions of new erythrocytes.
Introduction
The phylum Apicomplexa includes a large number of notorious human and animal pathogens such as Plasmodium falciparum, the causative agent of severe cases of malaria, killing millions of people every year and reported to be clinically responsible for death since Pharaon times.1 Host cell invasion by these obligate intracellular parasites occurs by active penetration of the host cell with the formation of a parasitophorous vacuole (PV). All along the erythrocytic cycle, and to achieve the needs of their growth, multiplication, and final release, Plasmodium parasites highly modify both the host cell membrane and their PV. Newly formed invasive parasites then escape from the host cell after the sequential opening of the PV and the host RBC membranes.2 Although some parasite proteins, and particularly a cascade of proteolytic activities,3 have been shown to be implicated, the dynamics of merozoite release, the crucial and very last step of the parasite intraerythrocytic development, and its mechanisms still remain to be deciphered.4 It is still commonly referred to as an “explosive” event,2,5,6 35 years after the seminal video microscopy work of Dvorak et al.7 However, what happens to the membrane during this “explosion” and, in particular, how parasite displacements can reach several times the parasite body size in a split second without any swimming appendices or inertia are questions that have not yet received convincing answers because of the lack of direct observations. Hypothetical mechanisms have been proposed but not proven, such as the shredding of the membrane because of the osmotic pressure or a breaching of the erythrocyte membrane by the parasite.5 Using high-speed video-microscopy, we have captured 41 individual events and identified new steps of merozoite egress showing that their release is indeed the result of an elastic instability of the infected RBC (iRBC) membrane that we relate to a high spontaneous curvature induced by the parasite.
Methods
Parasite culture
The P falciparum 3D7 strain was grown in vitro using human RBCs as described by Trager and Jensen8 in RPMI 1640 medium with N-glutamine supplemented with 25mM HEPES, 0.5% albumax I, 100μM hypoxanthine (Sigma-Aldrich), and 10 μg/mL gentamicin (Invitrogen), referred to as complete medium. The strain was adapted for optimal growth under 5% CO2 atmosphere at 37°C. Mature forms of P falciparum–iRBCs were enriched up to 80%-90% by gelatin flotation9 using gelofusin (Braun Medicals) and washed once with complete medium. iRBCs (2 × 108) suspended in 3 mL of complete medium were introduced into the observation chamber.
PKH labeling
Gelatin-enriched iRBCs (2 × 108) were labeled with PKH26 at 1μM final concentration, according to the manufacturer's protocol (Sigma-Aldrich). Labeled iRBCs were washed twice in complete medium and suspended in 3 mL of complete medium.
Observation protocol
The iRBC suspension is then injected into a chamber of observation. The chamber has a total volume of 3 mL and is composed of a top and a bottom separated by a Teflon spacer to fix the distance between the 2 glass slides to 500 μm. A continuous flow of 5% CO2 humidified gas is used to maintain the pH. The immersion objective and condenser are temperature-controlled to maintain the sample temperature to 37°C on an area of ∼ 1 cm2.
Using differential interference contrast (DIC) microscopy, we searched for a mature schizont and started recording at high enough frame rate until egress occurs. If egress did not happen within an hour, we focused on another mature schizont. For PKH26-labeled iRBCs, the protocol of recording was modified to limit photo-damage artefacts. Thus, DIC light was switched off and fluorescence illumination switched on only at the moment of egress. The record was only kept if egress happened in < 5 seconds. To exclude that the membrane events recorded were because of RBC photo damage, we have verified that healthy RBCs were not affected by fluorescence illumination during 30-second–long shots.
Both DIC and fluorescence microscopy observations were performed on a Leica DMIRB equipped with a 100× objective (numerical aperture [NA] 1.3) coupled to a high numerical aperture condenser (NA 1.3). The high-speed imaging was performed using a high-speed camera Phantom V7 (Vision Research Inc). Image analysis was performed using ImageJ (Freeware; National Institutes of Health) for image contrasting and filtering. The edge of the pore and the center of mass of each merozoite were tracked manually.
In total, 41 records were kept, of which 38 were representative of the elastic release we are describing, 2 were abortive egress events, and 1 remained unclear. Six DIC records and 4 fluorescence records happened in the focal plane allowing tracking of the rim radius evolution during RBC opening.
Results
Concordant with previously reported results, we have observed that merozoites are released after a swelling phase where the P falciparum iRBC becomes spherical and tensed2,6,7 (supplemental Video 1, available on the Blood Web site; see the Supplemental Materials link at the top of the online article). Using high-speed video microscopy, we have measured a total time scale of the release centered around 800 milliseconds and sometimes as fast as 400 milliseconds. The egress can be separated in 3 distinct steps as following. The initial step that we refer to as an “osmotic release” consists in the fast release of usually 1 and possibly 2 merozoites in 100-200 milliseconds, thus representing only a small fraction of the total time of egress. The 2 following successive steps occur on a larger time scale and correspond to a process that we call the “elastic release” of the left-behind merozoites, at distances of 10 μm, much larger than their own body size (estimated at 1.5 μm). We have further analyzed these newly observed steps by following and recording > 50 individual events of full merozoite release.
Osmotic release
As shown in Figure 1, the first merozoite is rapidly liberated with maximum velocities Vmax ranging from 5 to 110 μm/s (Figure 1B, see Video 1). As measured for 16 individual events of egress, Vmax shows a slight correlation with the initial radius R of the iRBC just before rupture: the smaller the iRBC, the faster the first released merozoite. It is noteworthy that we did not observe any significant difference in the number of merozoites per iRBC depending on the initial radius of the RBC.

Egress of the first merozoite. (A) Snapshots of the osmotically triggered ejection of the first merozoite using DIC microscopy. Scale bar, 3 μm. (B) Maximum velocity Vmax of the first merozoite as a function of the initial iRBC radius R, before membrane rupture. Error bars represent the error in the determination of the diameter of the iRBC before rupture.
Egress of the first merozoite. (A) Snapshots of the osmotically triggered ejection of the first merozoite using DIC microscopy. Scale bar, 3 μm. (B) Maximum velocity Vmax of the first merozoite as a function of the initial iRBC radius R, before membrane rupture. Error bars represent the error in the determination of the diameter of the iRBC before rupture.
Because the associated regime of pore opening is hampered by the release of the first parasite when using DIC microscopy and to monitor correctly this event, we have labeled the iRBC membrane using PKH26 and epifluorescence. As represented in Figure 2A, we clearly observe the initial regime of circular pore opening (supplemental Video 2; the opening of a pore in the iRBC, leading to the release of the first merozoite, is also observed in Figure 3 and supplemental Video 4). We report in Figure 2C, the values of the radius r of the pore as a function of time. These data clearly identify a change of regime in the RBC opening. Indeed, the curvature of the iRBC membrane changes when the pore radius is around r0 ≈ 1 μm. At this point, a shoulder-type deformation appears at the border of the growing hole as presented in Figure 2A (time 150 milliseconds compared with 100 milliseconds), before membrane curling starts.

iRBC opening regimes. (A) Sequence of images of PKH26-labeled iRBC using fluorescence microscopy and showing: the initial iRBCs (0 milliseconds), the circular pore opening (100 milliseconds), the shoulder type deformation of the RBC membrane (150 milliseconds) followed by the membrane curling (283 milliseconds). Scale bar, 1 μm. (B) Sequence of images of the first curl after the shoulder-type deformation (time lapse: 14.3 milliseconds). Scale bar, 1 μm. (C) Kinetics of the pore opening with the radius r of the pore as a function of time. Two regimes are identified: the circular opening and the curling taking place at t0. The parameters used for the data analysis are represented both in the inset and on the curve: the pore radius when curling starts r0 and the cell radius R. At any given time t, the opening is described by a pore radius r and a rim radius L. Error bars represent the error in the determination of the diameter of the pore.
iRBC opening regimes. (A) Sequence of images of PKH26-labeled iRBC using fluorescence microscopy and showing: the initial iRBCs (0 milliseconds), the circular pore opening (100 milliseconds), the shoulder type deformation of the RBC membrane (150 milliseconds) followed by the membrane curling (283 milliseconds). Scale bar, 1 μm. (B) Sequence of images of the first curl after the shoulder-type deformation (time lapse: 14.3 milliseconds). Scale bar, 1 μm. (C) Kinetics of the pore opening with the radius r of the pore as a function of time. Two regimes are identified: the circular opening and the curling taking place at t0. The parameters used for the data analysis are represented both in the inset and on the curve: the pore radius when curling starts r0 and the cell radius R. At any given time t, the opening is described by a pore radius r and a rim radius L. Error bars represent the error in the determination of the diameter of the pore.

Identification of 3 main steps in merozoite egress. (A) Snapshots of the whole release process: pressure driven ejection of the first parasite (up to 148 milliseconds); curling of the RBC membrane (192-324 milliseconds) and final buckling of the membrane (324-368 milliseconds) pushing the remaining merozoites forwards, far from their initial position. (B) Velocities as a function of time for the first merozoite (T; blue line), the parasite pigment (H; black line), the deepest merozoite at the bottom (B; green line), and the lowest point of the RBC membrane (M; red line).
Identification of 3 main steps in merozoite egress. (A) Snapshots of the whole release process: pressure driven ejection of the first parasite (up to 148 milliseconds); curling of the RBC membrane (192-324 milliseconds) and final buckling of the membrane (324-368 milliseconds) pushing the remaining merozoites forwards, far from their initial position. (B) Velocities as a function of time for the first merozoite (T; blue line), the parasite pigment (H; black line), the deepest merozoite at the bottom (B; green line), and the lowest point of the RBC membrane (M; red line).
Elastic release: curling and buckling
After the first ejection, the edge of the pore curls out and wraps into a toroidal rim whose major radius r grows in time (supplemental Videos 2 and 3; Figures 2 and 4A). This curling process was confirmed by direct observation of the first turn as presented in Figure 2B and supplemental Video 4. Then, when r reaches a critical value close to the radius R of the iRBC, the membrane changes the sign of its curvature (ie, from concave to convex) and buckles, liberating and dispersing the remaining merozoites as shown in Figure 4A. In Figure 4B, the velocities of the first top (T) and last bottom (B) merozoites as well as the malaria pigment hemozoin (H) and the extreme point (M) of the RBC membrane are plotted as a function of time. The curves indicate the separation between the time scales of the osmotic release of the first merozoite and the elastic release of the remaining merozoites. In Figure 4B, while the first released merozoite (T) is almost at rest, (H), (B), and (M) accelerate, with a velocity of the last released merozoites (B) almost as high as the velocity of the first one (T), ejected by the Laplace overpressure triggered by the osmotic stress. The high correlation between the velocities of (B) and (M) shows that the membrane elastic eversion is accompanying the motion of the parasites, transmitting momentum to the remaining merozoites more than ∼ 300 milliseconds. Moreover, the rapid change of velocity regime observed after 300 milliseconds characterizes the elastic instability of buckling driving the last regime of movement of the membrane at an increasing speed. This last step induces the release of the remaining merozoites with a wide angular distribution as shown on the last image of Figure 4A.

Efficient egress depends on the completed curling and buckling of the RBC membrane. Snapshots of iRBC aborted opening without buckling. The iRBC has been labeled using 1μM PKH26. The different steps of release are presented: pore opening (0.5 seconds), shoulder-type deformation (1 second) before curling. Curling stopped at 2 seconds, leaving merozoites trapped inside the cell (22 seconds).
Efficient egress depends on the completed curling and buckling of the RBC membrane. Snapshots of iRBC aborted opening without buckling. The iRBC has been labeled using 1μM PKH26. The different steps of release are presented: pore opening (0.5 seconds), shoulder-type deformation (1 second) before curling. Curling stopped at 2 seconds, leaving merozoites trapped inside the cell (22 seconds).
Thus, after the iRBC membrane curling, the release of the left-behind merozoites occurs together with the buckling of the membrane. In Figure 5, we show a sequence of images of an aborted egress, where most of the merozoites were trapped in the iRBC membrane because of an early stop of curling before any buckling of the membrane could occur (supplemental Video 5). These data strongly suggest that buckling is necessary for efficient release and dispersal of the merozoites.

Egress of merozoites from adhering versus nonadhering iRBCs. (A) Snapshots of merozoite release when the iRBC is adhering to the substrate. (B) Maximum velocity of the successively ejected merozoites as a function of time in the case of no adhesion (○) or adhesion (●). T, H, B, and M are defined in the legend of Figure 4B. Inset, The initial position of the iRBC before opening and the merozoite dispersion after RBC membrane buckling in the case of no adhesion and adhesion of the iRBC to the glass substrate. Error bars represent the scale of velocity for each individual merozoite because of errors in determining the position of its center of mass. Scale bar, 3 μm.
Egress of merozoites from adhering versus nonadhering iRBCs. (A) Snapshots of merozoite release when the iRBC is adhering to the substrate. (B) Maximum velocity of the successively ejected merozoites as a function of time in the case of no adhesion (○) or adhesion (●). T, H, B, and M are defined in the legend of Figure 4B. Inset, The initial position of the iRBC before opening and the merozoite dispersion after RBC membrane buckling in the case of no adhesion and adhesion of the iRBC to the glass substrate. Error bars represent the scale of velocity for each individual merozoite because of errors in determining the position of its center of mass. Scale bar, 3 μm.
Elastic release of merozoites from adhering infected erythrocytes
It is noteworthy that the data presented have been obtained with iRBCs in suspension and it is important to determine whether merozoite release proceeds through similar steps in vivo, when most red cells with mature parasites are sequestered in the microvasculature, adhering to endothelial cells. The observation of iRBCs adhered to the glass substrate shed some light on this process. Importantly, as presented in Figure 3 (supplemental Video 6), the same 3 steps of egress (pore opening, curling, and buckling) are observed but with some differences: we observe the sequential release of more than 5 merozoites through a r ≈ 1 μm pore with higher velocity compared with nonadhering iRBCs, before curling starts (Figure 3B). The process ends up with the projection of the membrane backward thus releasing merozoites but without actually pushing them forward (see Figure 3B inset).
Merozoite egress depends on a parasite-induced spontaneous curvature of the iRBC membrane
Polymersomes are artificial-sensitive copolymer-based vesicles10 and good candidates for drug carriers. Mabrouk et al have studied their bursting by a process showing striking similarities with the destabilization of the iRBC membrane leading to merozoite egress.11 Mabrouk et al have reported that the curling is driven by a molecularly imposed curvature c0 of the membrane opposite to and much higher than the initial curvature 1/R of the polymersome of radius R. According to the model by Mabrouk et al, once a pore nucleates, it grows because of the outer curling of the membrane releasing bending energy at a rate Pe. The dynamics of the rim is then controlled solely by the balance between Pe, and the viscous dissipation Pv because of the movement of the growing rim in the outer medium. We propose to describe the iRBC bursting with the same approach considering that at one point during the parasite development the iRBC membrane acquires a spontaneous curvature c0 different from a normal RBC and whose origin is yet to be determined. For iRBCs, however, the pore nucleation is immediately followed by the circular opening up to a radius r0, reached at a time t0. Details of our model modified from Mabrouk et al are provided in the supplemental data. The thickness e ≈ 50 nm of the plasma membrane has been determined using ultrasensitive force probe techniques on healthy RBCs.12 Using this value and that of the critical radius rc established from our data (Table 1), one can deduce the average spontaneous curvature c0 ≈ 21.0 ± 2.4 μm−1 of the membrane for nonlabeled iRBCs and c0 ≈ 11.5 ± 2.4 μm−1 for PKH26-labeled iRBCs, corresponding to radii of curvature of 48 and 87 nm, respectively. We have also measured the maximum pore radius rmax as well as the rim size Lmax before buckling as reported in Table 1. Using the values of c0 and rc and the expression of the rim radius L, one can compute L(rmax) ≈ 0.2 μm which is close to the measured values reported in Table 1.
Measured and fitted parameters of iRBC membrane curling
| R, μm | r0, μm | t0, s | rmax, μm | Lmax, μm | rc, μm | D, μm2/s |
|---|---|---|---|---|---|---|
| DIC | ||||||
| 3.1 | 1 | 0.7 | 2.6 | 0.35 | 0.3 ± 0.2 | 32 ± 3 |
| 3.1 | 1.4 | 0.15 | 3.1 | 0.3 | 0.3 ± 0.2 | 43 ± 2 |
| 3.0 | 0.9 | 0.45 | 2.7 | 0.5 | 0.4 ± 0.1 | 20 ± 1 |
| 2.7 | 0.5 | 0.01 | 2.6 | 0.4 | 0.3 ± 0.15 | 16 ± 1.5 |
| 2.9 | 1.5 | 0.25 | 3.1 | 0.4 | 0.2 ± 0.1 | 11 ± 1.5 |
| 3.0 | 1.1 | 0.06 | 2.6 | 0.4 | 0.3 ± 0.15 | 32 ± 3 |
| EpiFluo | ||||||
| 2.7 | 1 | 0.2 | 2.2 | 0.42 | 1.7 ± 0.6 | 36 ± 7.5 |
| 2.5 | 1.1 | 0.13 | 2.4 | 0.42 | 1 ± 0.2 | 41 ± 4 |
| 2.7 | 0.9 | 0.15 | 2.5 | 0.44 | 0.6 ± 0.6 | 44 ± 21 |
| 2.8 | 1.5 | 0.55 | 2.5 | 0.47 | 1 ± 0.25 | 12 ± 2 |
| R, μm | r0, μm | t0, s | rmax, μm | Lmax, μm | rc, μm | D, μm2/s |
|---|---|---|---|---|---|---|
| DIC | ||||||
| 3.1 | 1 | 0.7 | 2.6 | 0.35 | 0.3 ± 0.2 | 32 ± 3 |
| 3.1 | 1.4 | 0.15 | 3.1 | 0.3 | 0.3 ± 0.2 | 43 ± 2 |
| 3.0 | 0.9 | 0.45 | 2.7 | 0.5 | 0.4 ± 0.1 | 20 ± 1 |
| 2.7 | 0.5 | 0.01 | 2.6 | 0.4 | 0.3 ± 0.15 | 16 ± 1.5 |
| 2.9 | 1.5 | 0.25 | 3.1 | 0.4 | 0.2 ± 0.1 | 11 ± 1.5 |
| 3.0 | 1.1 | 0.06 | 2.6 | 0.4 | 0.3 ± 0.15 | 32 ± 3 |
| EpiFluo | ||||||
| 2.7 | 1 | 0.2 | 2.2 | 0.42 | 1.7 ± 0.6 | 36 ± 7.5 |
| 2.5 | 1.1 | 0.13 | 2.4 | 0.42 | 1 ± 0.2 | 41 ± 4 |
| 2.7 | 0.9 | 0.15 | 2.5 | 0.44 | 0.6 ± 0.6 | 44 ± 21 |
| 2.8 | 1.5 | 0.55 | 2.5 | 0.47 | 1 ± 0.25 | 12 ± 2 |
Values of rc and D (defined in the text) are obtained with the fit of the curves of the pore radius r as a function of time using equation 1 for iRBCs of radius R. Measured values t0 and r0 set the beginning of curling, and rmax and Lmax correspond, respectively, to the pore and rim radii before buckling. Two sets of data are presented for iRBCs without (DIC) and with (EpiFluo) PKH26 labeling. SE values of the parameters of the fit are indicated.
Discussion
We have video-recorded more than 50 individual events of malaria merozoite release from erythrocytes. Forty-one records were kept of which 2 were abortive events, 1 remained unclear and 38 allowed us to describe new steps of merozoite egress: osmotic release, pore opening in the red cell membrane allowing 1 merozoite, possibly 2, to be released and finally the curling and buckling of the erythrocyte membrane pushing free the remaining merozoites.
The osmotic origin of merozoite release has been previously proposed by Glushakova et al2 based on invasion efficiency measurements showing that hypertonic stress applied to iRBCs inhibited merozoite release, while hypotonic stress increased iRBC rupture. The correlation between the velocity of the first released merozoite and the radius of the iRBC strengthens the hypothesis of an osmotic origin of this release. Thus, the first merozoite release would result from an excess pressure in the inner liquid which is produced by the mechanical tension stored in the iRBC membrane during the initial osmotic swelling and is given by the Laplace law ΔP = 2σ0/R, where σ0 is the surface tension of the membrane before rupture and R the radius of the inflated iRBC. Once the pore opens, the pressurized liquid leaks out dragging out the first merozoite at a velocity Vmax likely of the order of magnitude of the flow velocity at the center of the pore of radius r, and given roughly by a pressure and viscous stress balance13 : ηVmax/r = ΔP(r) = 2σ(r)/R, where η is the viscosity of the outer fluid. The measurement of the velocity plotted in Figure 1 gives therefore an order of magnitude of the surface tension of the membrane at the egress of the very first parasite, considering that the pore radius is of the order of the ejected merozoite radius (0.5μm): σ(r) ∼ ηRVmax/r ∼ 6 × 10−8 − 6 × 10−7 N/m. Following Sandre et al, we assume that the surface tension decreases linearly with the surface of the pore leading to: σ(r) = σ0(1 − r2/rrelax2).13 rrelax represents the radius for which the membrane tension is fully relaxed and is given by Sandre et al: rrelax2 = 4R2σ0/KA, where KA is the expansion modulus for the RBC membrane whose value is dominated by the behavior of the lipid bilayer.13,14 Therefore, one can estimate the initial tension at the moment of rupture: σ0 = σ(r) + r2KA/(4R2) ≈ 2-3.5 mN/m, taking KA ≈ 300-500 mN/m.14 It is noteworthy that the tensed spherical shape of the iRBC is maintained on short time scales (< 30 seconds) before rupture. However, to occur within such a short time, the lysis of healthy RBC membrane needs tensions as high as 10-12 mN/m as obtained by micropipette aspiration techniques12,15 or by atomic force microscopy (AFM).16 Thus, the initial tension of rupture of an infected RBC membrane is lower than that of a healthy RBC, suggesting that the iRBC membrane has been weakened by the parasite at the time of release.17 Therefore, the lower membrane tension at the time of rupture might be because of modifications of the RBC membrane induced by the parasite and generating pore nucleation sites of lower energy. Obviously, it might be a heterogeneous process that occurs at preferential sites. One possible origin for these nucleation sites of lower energy is the clustering of band 3 triggered by its parasite-induced hyperphosphorylation.18
As defined by the release through a 1 μm pore, osmotic release is only observed for 1 and less frequently 2 merozoites when iRBC are in suspension and up to 5 merozoites for adhering iRBCs. These results suggest that adhesion maintains a membrane tension high enough to produce the overpressure driving more merozoites out of the iRBC.
However, although in agreement with the merozoite osmotic release proposed by Glushakova et al, our results clearly show that the build-up of osmotic pressure is not sufficient for the release of all merozoites; hence, full merozoite release needs another mechanism.2 We show here that in culture conditions, fully efficient release of the merozoites needs (1) additional opening of the iRBC membrane through curling and (2) dispersal of the remaining merozoites by buckling of the iRBC membrane. These processes were observed for iRBCs both in suspension and adhering to the glass substrate but with a significant difference on the parasite dispersion: for iRBCs in suspension, the release of merozoites by buckling of the RBC membrane leads to full egress and with a wider angular dispersal than for adhering iRBCs. Of note, the forward movement one can notice on supplemental Videos 1 and 3 is the result of the viscous drag encountered by the rim during curling and buckling. This eversion propels the entire system forward the same way one breaststroke propels a swimmer forward. If there was no drag, the center of mass of the system will not be moving. In vivo, iRBC are adhering to the endothelium of flowing vessels and the main role of eversion of the RBC membrane might simply be permissive in allowing flow dispersal unhindered by residual host-membrane restraints. In addition, the release of the first 5 merozoites with higher velocity might increase their efficiency of meeting, binding to and invading red cells in their close vicinity, particularly when iRBCs are involved in rosettes with uninfected RBCs.
The striking similarities observed between the destabilization of the iRBC membrane and the bursting of artificial-sensitive copolymer-based vesicles, or polymersomes,10 reported by Mabrouk et al,11 suggest that a common elastic mechanism is involved in both systems. We propose a model modified from Mabrouk et al and describing the iRBC bursting with the same approach (see supplemental data). Briefly, the balance between elastic power and viscous dissipation gives the law of variation of the radius r of the pore as a function of time:
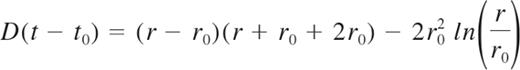
For time scales close to t0, equation 1 shows that r − r0 varies linearly in time with a slope equal to D/(2rc), while for larger time scales, or equivalently, when r is much larger compared with r0 and rc, (r − r0)2 ∼ D(t − t0) indicating a slower and diffusive-like opening of the pore. rc, which is equal to 2π/ec02 therefore represents the length scale of transition from the linear to the diffusive regime after the time t0. D, which is equal to κln/2eη, has the dimensions of a diffusion coefficient. As presented in Figure 2C, we have measured the variations of r as a function of time for individual events of iRBC opening and fitted our data using equation 1 to determine the values of rc and D reported in Table 1. This allowed us to determine the average spontaneous curvature c0 ≈ 21.0 ± 2.4 μm−1 of the membrane for nonlabeled RBCs and c0 ≈ 11.5 ± 2.4 μm−1 for PKH26-labeled RBCs. These spontaneous iRBC membrane curvatures correspond to radii of curvature of 48 and 87 nm, respectively, values close to the membrane thickness. In addition, we computed the rim radius L (see supplemental data) at rmax using the parameters rc and D and we found a value of 0.2 μm, which is close to the measured values Lmax reported in Table 1. Based on these observations, the number of turns before buckling is ∼ 4. Taken together, our results confirm that curling is because of a spontaneous curvature c0 acquired by the iRBC membrane. Subsequently, c0 should be sensitive to modifications of the lipid contents of the iRBC membrane and their relative repartition between the 2 membrane leaflets. Indeed, we observe a decrease of c0 measured for PKH26-labeled cells whose outer leaflet has been enriched by these extra lipids (Table 1). In addition cone-shape photo-damaged lipids might also account for the decrease of c0.
However, though the modified model fits the data and gives a reasonable value for the spontaneous curvature, it fails to predict the value obtained experimentally for D depending mainly on the membrane thickness and the viscosity. Using the typical value for the RBC membrane bending modulus κ ≈ 50 kBT,19 the viscosity of the outer medium close to that of water (η ≈ 10-3 Pa.s) and the membrane thickness e from 25 to 50 nm, we obtain D ≈ 4000-8000 μm2/s, that is values 2 orders of magnitude larger than the one reported in Table 1. The lowest value of e corresponds to a compact mesh of spectrin backed by the hardcore structure of transmembrane proteins, the glycocalix and the lipid bilayer,12,16 while the second value takes into account a thermally expanded structure of the spectrin network made of loops.12 Because of such a discrepancy between the fitted and the calculated D values, 2 assumptions of the model have to be reconsidered.
First, the model was based on a compact curling of the membrane. This hypothesis can be easily abandoned based on steric and dynamical arguments. Indeed, the compact curling might be hampered by structures remaining attached to the iRBC membrane after egress such as the submembrane skeleton and the Maurer clefts, previously shown to remain attached to the RBC membrane after egress.20-23 Moreover, studying the curling in a viscous fluid of an elastic stripe, with a spontaneous curvature, we observed that viscous friction prevents a compact curling as shown and discussed in the supplemental Figure 1A. The full description of such experiments is out of the scope of this article and this process is currently further investigated. However, our preliminary study shows that the interlayer distance between each curl is a fraction of c0-1 in the regime of flow for iRBC membrane curling (supplemental Figure 1B). Of note, electron micrographs of similar noncompact curls have been observed on the hypotonic lysis on ice-cold solutions of healthy RBCs fixed with OsO424 and corroborate the fact that dynamical curling is likely non compact with an interlayer distance of the order of c0-1. Such a correction for the effective value of the curling interlayer thickness is nevertheless not enough to explain the 2 order of magnitude of difference we obtain experimentally for D.
Thus, additional dissipative processes described by an effective viscosity ηeff 2 orders of magnitude larger than η would account for such a discrepancy. For instance, another possible source of extraviscous dissipation could come from the 2-dimensional (2D) membrane flow produced by the curling deformation. However it is not clear how to correctly calculate such a complex flow with a simple term in the energy balance described earlier. One crude approach to estimate such a 2D dissipation has been suggested in the supplementary files of Mabrouk et al.11 The authors considered that the viscous dissipation around the curling membrane edge could be represented by the same term as for a circular pore opening.13 After some algebra (supplemental data), it is easy to show that D depends mainly on the ratio ηS/ηr, where ηS is the surface viscosity of the membrane. Viscosity measurements have been done using many different techniques such as labeled molecular probe diffusion in bilayers,25 falling ball viscosimetry,26 phase-separated lipid domain diffusion,27 and pore-opening measurement techniques,28 establishing that ηS is ranging from 10-9 N.s/m. to 3.10−7 N.s/m. Consequently, D can vary by a factor 100, depending on the type of membrane flow. This parameter could thus explain the discrepancy between model-predicted and experimental D values.
Moreover, the lipid membrane of a RBC, although fluid like the membrane of a polymersome, is linked to a 2D submembrane spectrin network. Importantly, the parasite induces modifications of the RBC membrane and submembrane skeleton and links the Maurer clefts to the iRBC membrane from < 2 hours postinvasion up to merozoite release.20-23 Therefore, efficient egress of the merozoites might involve the release of interactions between the lipid membrane and these underlying structures.29 Thus, unzipping the RBC membrane from these structures could account for such a high effective viscosity ηeff and proteolytic activities reported to be crucial for release3,30-32 might be implicated. It is noteworthy that we have observed membrane fragments attached to the inner part of the curling membrane (supplemental Video 3) that could result from such unzipping activities and be the earlier stage of the blebs described on longer time scales by Glushakova et al.2 The role of the terminal state of the submembrane skeleton in the curling of the iRBC membrane has to be analyzed under the light of a recent 2D theoretical approach.33 Kabaso et al propose that a cleaved spectrin network remaining attached to the membrane induces intrinsic membrane curvature and generate curling, but on time scale 2 orders of magnitude higher than our observations. This approach is based on transmission electron microscopy micrographs24 showing the eversion of healthy RBC membrane, produced through profound hypotonic shocks in ice-cold media free of divalent cations. These ions are known to play a role on the release of the spectrin-actin bond.34 The generation of a curled state in such different conditions suggests that cleavage of the iRBC submembrane skeleton might participate in inducing the spontaneous curvature responsible for malaria merozoite egress.
Alternatively, if the integrity of the submembrane skeleton was preserved during curling, the spectrin network will be submitted to large local deformations that might modify the energy balance and could also account for the low D value obtained from the detailed model. The consequences of such deformations in addition to bending are currently investigated.
Conclusion
This article describes for the first time the full process of the RBC membrane opening leading to the release and efficient dispersion of invasive malaria merozoites from cultured iRBCs. Contrary to previous descriptions, we show that osmotic release accounts only for the egress of few merozoites in a first step, while the subsequent curling and buckling of the iRBC membrane release all remaining merozoites. Importantly, similar steps occur for adhering iRBCs but with some differences. Our observations suggest that, in vivo, more merozoites might be propelled by the flow resulting from a sustained membrane tension; the final curling-eversion process thus exposes the remaining merozoites to the blood flow that might facilitate their dispersion. Our data strongly suggest that the modifications of the RBC membrane by the parasite and the contribution of parasite proteins to interactions between this host cell membrane and submembrane structures like the RBC skeleton and the Maurer clefts are central to this elastic release. The identification of the protein-protein interactions implicated needs further investigation and should provide new targets for antimalarial compounds. Moreover, our study sheds new light on pore opening in the RBC membrane and questions the role of the cytoskeleton and, more generally, the role of the different mechanical properties of the membrane, such as shear elasticity in membrane opening. Finally, the mechanisms of curling and buckling by which malaria merozoites egress from RBCs could be more general and bring some understanding of other biologic systems that might imply similar physical concepts.35
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
We thank V. Lorman and J. F. Dubremetz for helpful discussions and E. Vialla for technical assistance in parasite cultures. UMR 5587 belongs to the CNRS French Consortium “CellTiss.”
This work was supported by the French Ministry of Research, the CNRS Physics-Chemistry-Biology Interdisciplinary Program, and the University Montpellier 2 Interdisciplinary Program.
Authorship
Contribution: M.A., G.M., L.B., and M.R. performed experiments; M.A., G.M., and C.B.-B. designed the study, analyzed data, and wrote the paper; and all authors discussed the results and commented on the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Catherine Braun-Breton, University Montpellier II, CNRS UMR 5235, Dynamique des Interactions Membranaires Normales et Pathologiques, 34095 Montpellier cedex 5, France; e-mail: cbb@univ-montp2.fr.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal