

Key Points
Emperipolesis (neutrophil transit through megakaryocytes) occurs in fast and slow forms that differ morphologically.
Intramegakaryocytic neutrophils reside in emperisomes and in cytoplasm near the demarcation membrane system, endoplasmic reticulum, and nucleus.

Abstract
Neutrophils transit through megakaryocytes in a process termed emperipolesis, but it is unknown whether this interaction is a single type of cell-in-cell interaction or a set of distinct processes. Using a murine in vitro model, we characterized emperipolesis by live-cell spinning disk microscopy and electron microscopy. Approximately half of neutrophils exited the megakaryocyte rapidly, typically in 10 minutes or less, displaying ameboid morphology as they passed through the host cell (fast emperipolesis). The remaining neutrophils assumed a sessile morphology, most remaining within the megakaryocyte for at least 60 minutes (slow emperipolesis). These neutrophils typically localized near the megakaryocyte nucleus. By ultrastructural assessment, all internalized neutrophils remained morphologically intact. Most neutrophils resided within emperisomes, but some could be visualized exiting the emperisome to enter the cell cytoplasm. Neutrophils in the cytoplasm assumed close contact with the platelet-forming demarcation membrane system or the perinuclear endoplasmic reticulum. These findings reveal that megakaryocyte emperipolesis reflects at least 2 distinct processes differing in transit time and morphology, fast and slow emperipolesis, suggesting divergent physiologic functions.
Introduction
Megakaryocytes (MKs) are the largest cells in the bone marrow (50-100 µm) and constitute ∼0.05% of marrow cells.1 MKs produce platelets by extending long protrusions called proplatelets into sinusoids where shear stress causes platelet release into the circulation.2,3 This ability to generate platelets has been extensively studied. Recent observations have begun to suggest that MKs also have important immune functions.4,5 MKs express Toll-like receptors6-9 and other immune receptors10-12 and produce inflammatory cytokines and chemokines.13-15 Early MK progenitors express major histocompatibility complex (MHC) class II.16 Mature MKs cross-present antigens to CD8+ T cells via MHC I17 to CD4+ T cells via MHC II18 and exhibit antiviral potency.19 In COVID-19 patients, the percentage of MKs in the peripheral blood mononuclear cell fraction is increased, and a hyperinflammatory MK subset, enriched in critically ill individuals, may contribute to systemic inflammation.20 Thus, the functional portfolio of MKs extends considerably beyond platelet production.
One intriguing functional specialization of MKs is to interact directly with leukocytes, predominantly neutrophils, in a cell-in-cell interaction termed emperipolesis (EP).21 Derived from the Greek for “inside round about wandering,” EP was first described in 1956 by Humble et al.22 Passage through MKs occurs without apparent harm to either cell.23,24 Efficient neutrophil EP requires active cytoskeletal rearrangement by both the host MK and the transiting neutrophil.24 These features distinguish EP from cell-in-cell interactions such as phagocytosis or entosis in which the engulfed cell remains passive and is typically digested.25 Under physiological conditions, conventional paraffin sections identify EP in approximately 2% to 5% of MKs in healthy mice.24 This frequency can more than double with systemic inflammation,24 chronic blood loss,26 myelofibrosis,27-29 myeloproliferative diseases,30 and gray platelet syndrome.31-34
Although regularly observed, basic questions regarding the cell biology of EP remain unanswered. We showed previously that neutrophils undergoing EP can fuse transiently with the MK demarcation membrane system (DMS), thereby transferring neutrophil membrane to daughter platelets and enhancing platelet production.24 Earlier authors postulated that EP may serve as a transmegakaryocytic route for neutrophils in the bone marrow to enter the circulation26 or that MKs might provide a “sanctuary” for neutrophils.35 Because EP is observed in multiple states of health and disease, EP may serve correspondingly diverse functions.
We hypothesized that if EP represented a heterogeneous set of processes, then the transit of neutrophils through MKs could exhibit corresponding morphological heterogeneity. We therefore employed immunofluorescence and electron microscopy (EM) to investigate the fate of neutrophils engaged in EP. We demonstrate here that EP diverges into fast and slow forms, with multiple distinct intermediate stages, suggesting at least two processes with potentially divergent physiological roles.
Materials and methods
Mice
Eight- to 12-week-old wild-type C57BL/6J mice were purchased from the Jackson Laboratory (#000664) and housed at specific pathogen-free conditions. All animal studies were approved by the Institutional Animal Care and Use Committee of the Brigham and Women’s Hospital.
Antibodies and reagents
Anti-CD41 allophycocyanin (APC) (MWReg30), anti-CD41 AF488 (MWReg30), and anti-Ly6G AF594 (1A8) were from BioLegend. Polyclonal anticalnexin and FluorSave Reagent were from Sigma-Aldrich. Polyclonal anti-golgin-97, donkey anti-rabbit AF488, DRAQ5, Hoechst 33342, RPMI 1640 with and without phenol red, ammonium-chloride-potassium lysing buffer, paraformaldehyde, and glutaraldehyde were from Thermo Fisher.
Isolation of murine bone marrow cells
Femurs and tibias were flushed with phosphate-buffered saline (PBS) using 22-gauge needles. Cell suspensions were then filtered through 40 µm cell strainers to remove pieces of bone or tissue and centrifuged, followed by lysis of red blood cells using ammonium-chloride-potassium lysing buffer. Bone marrow cells were then washed with PBS and resuspended in complete RPMI medium supplemented with 1% supernatant from the thrombopoietin (TPO)-producing fibroblast cell line GP122 (hereafter called TPO medium).36
Isolation of murine megakaryocytes
Hematopoietic progenitor cells were isolated from bone marrow using the EasySep Mouse Hematopoietic Progenitor Cell Isolation Kit (negative selection) and cultured 1, 2, or 4 days in TPO medium (5 × 106 cells/mL). Alternatively, bone marrow cells were cultured in TPO medium (107 cells/mL) for 4 days at 37°C and 5% CO2. MKs were then enriched using an albumin step gradient.37
Isolation of murine marrow and blood neutrophils and peritoneal cells
Marrow cells were obtained as described above. Circulating cells were obtained via cardiac puncture followed by red blood cell lysis. Marrow and circulating neutrophils were isolated by negative depletion (Neutrophil Isolation Kit, Miltenyi). Neutrophils recruited to the inflamed peritoneum were obtained by washing the peritoneum with 5 mL cold PBS 2 hours after intraperitoneal (IP) injection of 25 ng IL-1β. Emperipolesis was performed with 2 × 104 MKs cocultured with 2 × 105 Ly6G+ neutrophils (marrow and blood) or 2 × 105 unsorted peritoneum cells.
Emperipolesis assay
2 × 106 bone marrow cells and 2 × 104 MKs were cocultured in P96 round-bottom wells for 12 hours at 37°C and 5% CO2.
Laser scanning confocal microscopy
After 12 hours of coculture, bone marrow cells and MKs were fixed in paraformaldehyde 2% for 30 minutes at room temperature. After washing with PBS, cells were resuspended in PBS containing 0.2% saponin and 10% fetal bovine serum (permeabilization buffer) and stained with Hoechst 33342 (5 µg/mL), anti-CD41 AF488, and anti-Ly6G AF594 for 4 hours at room temperature or overnight at 4°C. In some experiments, cells were stained with Hoechst 33342 (5 µg/mL), anti-CD41 APC, anti-Ly6G AF594, and anti-calnexin or anti-golgin-97 (2.5 µg/mL, respectively). Cells were then washed with PBS and resuspended in permeabilization buffer containing donkey anti-rabbit AF488 secondary antibody (10 µg/mL) for 4 hours at room temperature or overnight at 4°C. After staining, cells were washed and cytospun onto coverslips and mounted on glass microscope slides (Fisher Scientific) using FluorSave Reagent. Images were obtained using a Zeiss LSM 800 with Airyscan attached to a Zeiss Axio Observer Z1 Inverted Microscope using a Plan-Apochromat 63x objective. Zen 2.3 blue edition software was used for image acquisition. Image analysis was performed using ImageJ 1.52p.
Spinning disk confocal microscopy
Neutrophils and MKs were stained with anti-CD41 AF488 and anti-Ly6G AF594 (1.5 µg/mL, respectively) for 1 hour prior to the experiment. DNA was stained with Hoechst 33342 (5 µg/mL) or DRAQ5 (5 µM). Cells were resuspended in TPO medium without red phenol to minimize autofluorescence and plated onto Nunc Glass Bottom Dishes (Thermo Fisher) for imaging. Images were obtained using a W1 Yokogawa Spinning Disk Confocal attached to a Nikon Ti inverted microscope with a Plan Fluor 40x/1.3 Oil DIC H/N2 objective and Nikon Elements Acquisition Software AR 5.02 or using a Perkin Elmer Ultraview Vox Spinning Disk Confocal attached to a Nikon Ti inverted microscope with a 60x (1.4NA) objective and Volocity Acquisition Software 6.3. Microscopy chambers were kept at 37°C and 5% CO2 throughout the experiment. In each experiment, 3 regions of interest and 10 to 12 z-stacks were imaged, with approximately 90 seconds between 2 timepoints. Image analysis was performed using ImageJ 1.52p.
Tracking of neutrophil migration
The migration of neutrophils through the cytoplasm of MKs was tracked using the ImageJ plugin TrackMate v4.0.1.
Electron microscopy
After 12 hours of coculture, bone marrow cells and MKs were washed twice with PBS and fixed in paraformaldehyde 2% and glutaraldehyde 0.1% for 4 hours at room temperature. Specimens were postfixed in 1% osmium tetroxide and 1.5% potassium ferrocyanide, and stained with 1% uranyl acetate, followed by gradual dehydration in 70%, 90%, and 100% ethanol and propylene oxide. Specimens were then embedded in Epon. Eighty nanometer sections were imaged using a JEOL 1200EX transmission electron microscope. EM imaging was performed in the Harvard Medical School Electron Microscopy Facility.
Statistical analysis
Statistical analyses were performed using Graphpad Prism 8 or R. One-way ANOVA and post hoc Tukey test were performed to compare the frequency of EP across developmental stages. Hartigan’s dip test was performed to determine bimodality in transit time. Unpaired Student t test was performed to compare neutrophil migration speed between fast and slow EP. All values were displayed as mean ± standard error of the mean. A P value ≤.05 was considered statistically significant. *P ≤ .05, **P ≤ .01, ***P ≤ .001, ****P ≤ .0001.
Results
Neutrophil emperipolesis is most efficient in mature megakaryocytes
To study EP, we employed a model wherein murine MKs differentiated from hematopoietic progenitor cells are incubated together with unfractionated murine bone marrow. We performed EP with MKs at different stages in culture, corresponding to different maturational states, grading MK maturity using the extent of DMS.38 In each of 3 experiments, we evaluated EP as performed by 100 MKs at each maturational stage. Mature MKs proved most efficient at EP, displaying neutrophil uptake by ∼20% of cells in comparison with 0.33% of immature MKs (Figure 1A-G). Uptake of >1 neutrophil was largely restricted to the most mature MKs (Figure 1H-I). We therefore employed mature day-4 MK cultures for our studies going forward.
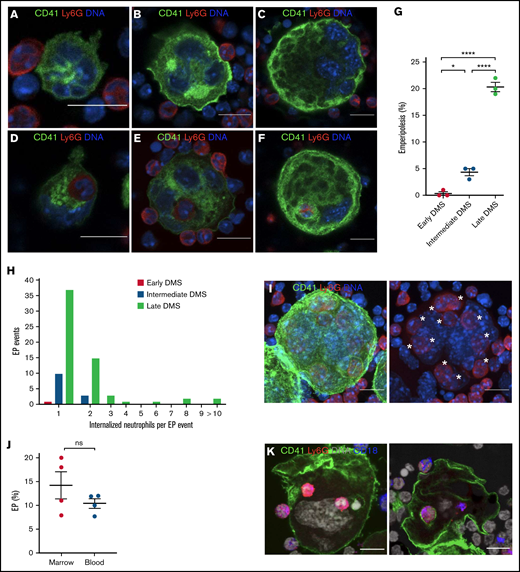
Visualization of EP in MKs of different maturational stages. Hematopoietic progenitor cells were cultured in TPO medium for 1, 2, or 4 days to obtain MKs of different maturation levels. MKs were then cocultured with bone marrow cells for 12 hours. MKs were stained with anti-CD41 AF488 (green), neutrophils were stained with anti-Ly6G AF594 (red), and DNA was stained with Hoechst 33342 (blue). MK maturity was graded based on the extent of the DMS (early, intermediate, and late DMS). Images were obtained using a Zeiss LSM 800 with Airyscan attached to a Zeiss Axio Observer Z1 Inverted Microscope with a Plan-Apochromat 63x objective. Scale bars, 10 µm. (A) Immature MK showing DMS beginning to develop between the nuclear lobes and forming connections with the MK surface (early DMS). (B) With increasing maturation, the DMS becomes more prominent and forms thicker connections with the MK surface (intermediate DMS). (C) Mature MK with extensive DMS occupying the majority of the MK cytoplasm (late DMS). (D-F) Early, intermediate, and late DMS MKs engulfing neutrophils during EP. (G) EP frequency across MK maturational stages. 100 MKs per maturational stage per experiment were counted in each of 3 independent experiments. (H) The number of engulfed neutrophils per EP event across MK maturational stages. Pooled data from 3 independent experiments (n = 300 MKs per maturational stage; EP events: early DMS MKs: 1, intermediate DMS MKs: 13, and late DMS MKs: 61). (I) Z-projection of mature MK (late DMS) containing 11 neutrophils (*s). (J) EP assay with neutrophils isolated from blood or bone marrow; 4 independent experiments. (K) EP assay with peritoneal cells harvested 2 hours after IP injection of 25 ng/mL IL1B. Green, CD41; red, Ly6G; blue, CD18; gray, DNA. Scale bars, 10 µm.
Visualization of EP in MKs of different maturational stages. Hematopoietic progenitor cells were cultured in TPO medium for 1, 2, or 4 days to obtain MKs of different maturation levels. MKs were then cocultured with bone marrow cells for 12 hours. MKs were stained with anti-CD41 AF488 (green), neutrophils were stained with anti-Ly6G AF594 (red), and DNA was stained with Hoechst 33342 (blue). MK maturity was graded based on the extent of the DMS (early, intermediate, and late DMS). Images were obtained using a Zeiss LSM 800 with Airyscan attached to a Zeiss Axio Observer Z1 Inverted Microscope with a Plan-Apochromat 63x objective. Scale bars, 10 µm. (A) Immature MK showing DMS beginning to develop between the nuclear lobes and forming connections with the MK surface (early DMS). (B) With increasing maturation, the DMS becomes more prominent and forms thicker connections with the MK surface (intermediate DMS). (C) Mature MK with extensive DMS occupying the majority of the MK cytoplasm (late DMS). (D-F) Early, intermediate, and late DMS MKs engulfing neutrophils during EP. (G) EP frequency across MK maturational stages. 100 MKs per maturational stage per experiment were counted in each of 3 independent experiments. (H) The number of engulfed neutrophils per EP event across MK maturational stages. Pooled data from 3 independent experiments (n = 300 MKs per maturational stage; EP events: early DMS MKs: 1, intermediate DMS MKs: 13, and late DMS MKs: 61). (I) Z-projection of mature MK (late DMS) containing 11 neutrophils (*s). (J) EP assay with neutrophils isolated from blood or bone marrow; 4 independent experiments. (K) EP assay with peritoneal cells harvested 2 hours after IP injection of 25 ng/mL IL1B. Green, CD41; red, Ly6G; blue, CD18; gray, DNA. Scale bars, 10 µm.
Circulating blood neutrophils were approximately as efficient as bone marrow neutrophils for emperipolesis (Figure 1J). Neutrophils infiltrating into peritoneum inflamed by IP IL-1β were also capable of emperipolesis (Figure 1K), demonstrating that emperipolesis is not restricted to immature or resting neutrophils. Marrow neutrophils from mice treated IP with PBS or IL-1β entered MKs with equal efficiency (supplemental Figure 1), suggesting that the enhanced emperipolesis observed in inflamed mice in vivo24 reflects the impact of activation signals on the MKs themselves or other factors rather than a change intrinsic to neutrophils alone.
Because MKs are localized primarily within the marrow compartment, we chose to work with marrow neutrophils for the rest of this study.
Bimodal neutrophil transit time through megakaryocytes: fast and slow emperipolesis
To understand whether neutrophil transit through MKs is uniform or heterogeneous, we employed spinning disk confocal microscopy. MKs were cultured together with whole bone marrow cells and visualized over 90 minutes, obtaining images every 90 seconds and acquiring 10 to 12 z-stacks per MK to distinguish internalized from superimposed neutrophils. We analyzed 28 EP events. Half (14/28) of neutrophils completed passage through MKs in 30 minutes or less (Figure 2A-B; supplemental Video 1). Of these “fast emperipolesis” events, most (11/14; 79%) were completed within 10 minutes, with some episodes of EP requiring as little as 3 minutes. By contrast, other neutrophils resided within MKs for at least 50 minutes (13/28; 46%), with most (12/13; 92%) remaining for >60 minutes (“slow emperipolesis”) (Figure 2A,C; supplemental Video 2). Hartigan’s dip test39 confirmed a bimodal distribution of transit times (P = 7.89 × 10−7). Interestingly, fast and slow EP could be observed simultaneously in the same MK (Figure 3A). These findings show that experimental EP encompasses fast and slow forms with distinct time courses.
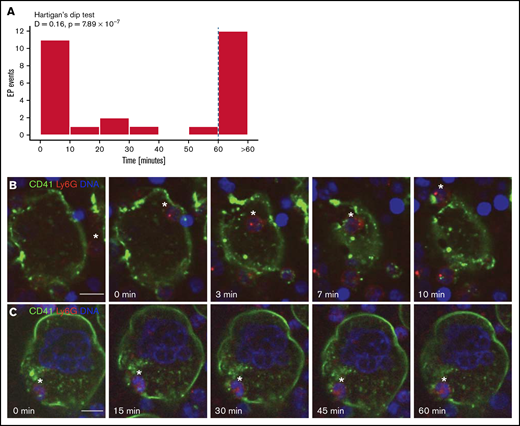
Neutrophil transit time through MKs is bimodally distributed: fast and slow EP. Mature MKs were stained with anti-CD41 AF488 (green) and coincubated with fresh bone marrow cells stained with anti-Ly6G AF594 (red). DNA was stained with DRAQ5 (blue). (A) Histogram depicting the duration of neutrophil transit through MKs of 28 EP events reveals a bimodal distribution with peaks between 0 and 10 minutes (fast EP) and >60 minutes (slow EP). Results pooled from 5 independent experiments. Bimodality was confirmed by Hartigan’s dip test (D = 0.16, P = 7.89 × 10−7). (B-C) Images were obtained using a W1 Yokogawa Spinning Disk Confocal attached to a Nikon Ti inverted microscope with a Plan Fluor 40x/1.3 Oil DIC H/N2 objective. Scale bars, 10 µm. (B) Representative image sequence of fast EP. The neutrophil (*) enters the MK on the right side, migrates through the MK cytoplasm, and egresses on the opposite side within 10 minutes. (C) Representative image sequence of slow EP. The neutrophil (*) is already inside the MK at the beginning of the image acquisition and remains inside for at least 60 minutes, showing no migration inside the MK.
Neutrophil transit time through MKs is bimodally distributed: fast and slow EP. Mature MKs were stained with anti-CD41 AF488 (green) and coincubated with fresh bone marrow cells stained with anti-Ly6G AF594 (red). DNA was stained with DRAQ5 (blue). (A) Histogram depicting the duration of neutrophil transit through MKs of 28 EP events reveals a bimodal distribution with peaks between 0 and 10 minutes (fast EP) and >60 minutes (slow EP). Results pooled from 5 independent experiments. Bimodality was confirmed by Hartigan’s dip test (D = 0.16, P = 7.89 × 10−7). (B-C) Images were obtained using a W1 Yokogawa Spinning Disk Confocal attached to a Nikon Ti inverted microscope with a Plan Fluor 40x/1.3 Oil DIC H/N2 objective. Scale bars, 10 µm. (B) Representative image sequence of fast EP. The neutrophil (*) enters the MK on the right side, migrates through the MK cytoplasm, and egresses on the opposite side within 10 minutes. (C) Representative image sequence of slow EP. The neutrophil (*) is already inside the MK at the beginning of the image acquisition and remains inside for at least 60 minutes, showing no migration inside the MK.
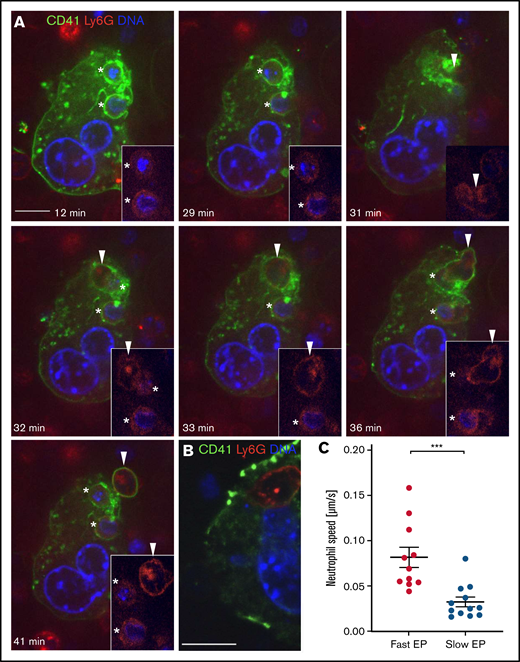
Distinct morphology of fast and slow EP within a single MK. Mature MKs were stained with anti-CD41 AF488 (green) and coincubated with fresh bone marrow cells stained with anti-Ly6G AF594 (red). DNA was stained with Hoechst 33342 (blue). (A-B) Images were obtained using a Perkin Elmer Ultraview Vox Spinning Disk Confocal attached to a Nikon Ti inverted microscope with a 60x (1.4NA) objective. Scale bars, 10 µm. (A) An MK showing fast and slow EP simultaneously to illustrate morphological differences of both forms. Two neutrophils undergoing slow EP (*) assume a sessile state. A third neutrophil (arrowhead) enters the MK after 31 minutes and extends dynamic membrane protrusions to propel itself through the MK cytoplasm, exiting within few minutes. (B) Representative image of a slow EP neutrophil residing near the MK nucleus. (C) The passage of neutrophils undergoing fast EP (<10 minutes) and slow EP (>60 minutes) through MKs was tracked using the ImageJ plugin TrackMate to determine the mean speed of migration.
Distinct morphology of fast and slow EP within a single MK. Mature MKs were stained with anti-CD41 AF488 (green) and coincubated with fresh bone marrow cells stained with anti-Ly6G AF594 (red). DNA was stained with Hoechst 33342 (blue). (A-B) Images were obtained using a Perkin Elmer Ultraview Vox Spinning Disk Confocal attached to a Nikon Ti inverted microscope with a 60x (1.4NA) objective. Scale bars, 10 µm. (A) An MK showing fast and slow EP simultaneously to illustrate morphological differences of both forms. Two neutrophils undergoing slow EP (*) assume a sessile state. A third neutrophil (arrowhead) enters the MK after 31 minutes and extends dynamic membrane protrusions to propel itself through the MK cytoplasm, exiting within few minutes. (B) Representative image of a slow EP neutrophil residing near the MK nucleus. (C) The passage of neutrophils undergoing fast EP (<10 minutes) and slow EP (>60 minutes) through MKs was tracked using the ImageJ plugin TrackMate to determine the mean speed of migration.
Distinct morphology of fast and slow emperipolesis
We characterized the morphological features of fast and slow EP. Neutrophils undergoing fast EP rapidly transited through MKs by extending dynamic membrane protrusions, appearing to propel themselves through the host cell (Figure 3A; supplemental Video 3). By contrast, in slow EP, resident neutrophils remained in a single location within the MK for the entire duration without any signs of mobility (Figure 3A; supplemental Video 3). These long-term resident neutrophils localized adjacent to the MK nucleus in 42% of events (5/12), in some cases showing particularly close approximation (Figure 3B). We then tracked neutrophil movement within MKs for fast and slow EP. Neutrophils undergoing fast EP showed a 2.5-fold higher mean migration speed inside MKs compared with neutrophils undergoing slow EP (Figure 3C). These results confirm our observation that slow EP neutrophils remain largely sessile. Fast and slow EP are thus morphologically as well as chronologically distinct.
Characterization of the ultrastructural features of emperipolesis by electron microscopy
Prior studies have shown that neutrophils engaged in EP may reside either in MK vacuoles, termed emperisomes, or directly within the MK cytoplasm.24 We sought to better understand the relationship between these compartments using transmission EM. We cocultured murine MKs and bone marrow cells as above and processed them for EM after 12 hours, analyzing 45 EP events across 5 independent experiments.
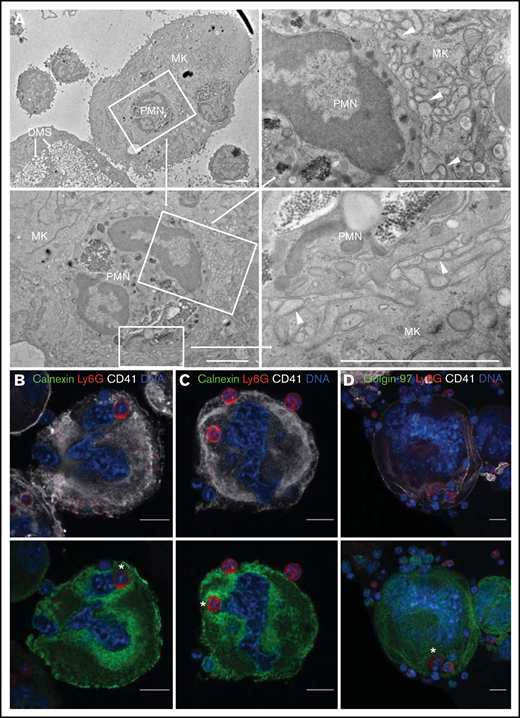
All MKs and neutrophils remained morphologically intact without membrane blebbing, nuclear fragmentation, or other evidence of apoptosis. Residence within an emperisome was the most common location for internalized neutrophils (18 of 45 events, 40%), yet the interaction between neutrophil and emperisome was heterogeneous. In some cases, the emperisome membrane was smooth and separated from the neutrophil by a large pericellular space (Figure 4Ai). Alternately, the emperisome membrane could exhibit small protrusions extending toward the engulfed neutrophil (Figure 4Aii). Finally, the emperisome could be tightly wrapped around the neutrophil, with zipper-like approximation of neutrophil and emperisome membranes (Figure 4Aiii). Intriguingly, some neutrophils appeared within a vacuolar space but also in contact with the cytoplasmic DMS without an interposed MK membrane, suggesting an egress event mediated by penetration through the emperisome (Figure 4Bi). Other neutrophils were entirely surrounded by DMS (Figure 4Bi). Importantly, some neutrophils could be visualized transiting directly from emperisome into the MK cytoplasm, far from the DMS (Figure 4Ci, including blowup images and 4Cii showing dual residence of a neutrophil in emperisome [Figure 4Cia] and cytoplasm [Figure 4Cib]), or fully resident within the MK cytoplasm without any interposed MK membrane (Figure 4Cii). The frequencies of these different EP stages are shown in Figure 4D. Of note, despite the proximity of the neutrophil to the MK nucleus in some instances, our EM studies identified no examples of direct contact between neutrophil and nuclear membranes. Although the fixed nature of EM images precludes assignment to fast or slow EP, these images confirm the highly varied interaction between an internalized neutrophil and its host MK and suggest that direct penetration through the emperisome membrane represents the most common mechanism of neutrophil egress into the MK cytoplasm.

Characterization of the ultrastructural features of EP by electron microscopy. Mature MKs incubated with bone marrow cells for 12 hours were fixed and processed for transmission EM. Forty-five EP events were observed from 5 experiments. (Ai-Cii) Transmission EM images of EP. Scale bars, 2 µm or (for magnification of Aiii) 500 nm. (Ai) Large, round emperisome with a smooth vacuolar membrane surrounding a neutrophil (polymorphonuclear leukocyte, PMN). (Aii) The emperisome extends membrane protrusions toward the engulfed neutrophil. (Aiii) The emperisome tightly wraps around the engulfed neutrophil. Magnification shows close membrane approximation between neutrophil and emperisome membranes (arrowheads). (Bi) Internalized neutrophil partly covered by the emperisome and partly exposed to the DMS of the MK. (Bii) Neutrophil residing within the cavities of the DMS. (Ci) Internalized neutrophil partly covered by the emperisome (Cia) and partly exposed to organelles of the MK cytoplasm (Cib). (Cii) Two neutrophils fully reside inside the MK cytoplasm. Only the neutrophil membranes remain visible (arrowheads). (D) Frequency of the previously described EP stages (n = 45).
Characterization of the ultrastructural features of EP by electron microscopy. Mature MKs incubated with bone marrow cells for 12 hours were fixed and processed for transmission EM. Forty-five EP events were observed from 5 experiments. (Ai-Cii) Transmission EM images of EP. Scale bars, 2 µm or (for magnification of Aiii) 500 nm. (Ai) Large, round emperisome with a smooth vacuolar membrane surrounding a neutrophil (polymorphonuclear leukocyte, PMN). (Aii) The emperisome extends membrane protrusions toward the engulfed neutrophil. (Aiii) The emperisome tightly wraps around the engulfed neutrophil. Magnification shows close membrane approximation between neutrophil and emperisome membranes (arrowheads). (Bi) Internalized neutrophil partly covered by the emperisome and partly exposed to the DMS of the MK. (Bii) Neutrophil residing within the cavities of the DMS. (Ci) Internalized neutrophil partly covered by the emperisome (Cia) and partly exposed to organelles of the MK cytoplasm (Cib). (Cii) Two neutrophils fully reside inside the MK cytoplasm. Only the neutrophil membranes remain visible (arrowheads). (D) Frequency of the previously described EP stages (n = 45).
Neutrophils interact with the megakaryocyte endoplasmic reticulum as well as the DMS
The DMS is easily recognized by its dilated appearance (Figure 5A, bottom left), but not all interactions between cytoplasmic neutrophils and MK organelles were with the DMS. Identifying membranes by EM as belonging to the endoplasmic reticulum (ER) or the Golgi apparatus can be difficult. Seven of 45 neutrophils (16%) were surrounded by membranes that we could not unambiguously assign to one of these structures (Figures 4D and 5A). Given our observation that neutrophils undergoing slow EP frequently reside near the MK nucleus, we hypothesized that they might interact with the perinuclear ER. Immunofluorescence staining confirmed that the perinuclear ER sometimes surrounded intracytoplasmic neutrophils (Figure 5B-C), enclosing neutrophils between ER and nucleus. This location is distinct from that of the DMS, as reflected in the inverse distribution of the calnexin+ ER and the CD41+ DMS (Figure 5B-C). The Golgi apparatus, another intracellular membrane network marked by golgin-97, did not colocalize with internalized neutrophils (Figure 5D). These data further confirm the diversity of intramegakaryocytic localization by neutrophils in EP, supporting the heterogeneity of this process.
Neutrophils interact with the MK endoplasmic reticulum as well as the DMS. MKs were allowed to engage in EP for 12 hours followed by processing for EM or laser scanning confocal microscopy. (A) Transmission EM images of EP. The internalized neutrophil is surrounded by a membrane network (arrowheads) that does not resemble the DMS. Scale bars, 2 µm. (B-D) Cells were stained with anti-CD41 APC (white), anti-Ly6G AF594 (red), anti-calnexin or anti-golgin-97 (green), and Hoechst 33342 (blue). Images were obtained using a Zeiss LSM 800 with Airyscan attached to a Zeiss Axio Observer Z1 Inverted Microscope with a Plan-Apochromat 63x objective. Scale bars, 10 µm. (B-C) The perinuclear portion of the MK endoplasmic reticulum (ER) surrounds the internalized neutrophils (*s). The neutrophils are localized between the MK nucleus and ER. Note the inverse distribution of the DMS and ER. (D) Internalized neutrophils (*) did not colocalize with the Golgi apparatus of MKs.
Neutrophils interact with the MK endoplasmic reticulum as well as the DMS. MKs were allowed to engage in EP for 12 hours followed by processing for EM or laser scanning confocal microscopy. (A) Transmission EM images of EP. The internalized neutrophil is surrounded by a membrane network (arrowheads) that does not resemble the DMS. Scale bars, 2 µm. (B-D) Cells were stained with anti-CD41 APC (white), anti-Ly6G AF594 (red), anti-calnexin or anti-golgin-97 (green), and Hoechst 33342 (blue). Images were obtained using a Zeiss LSM 800 with Airyscan attached to a Zeiss Axio Observer Z1 Inverted Microscope with a Plan-Apochromat 63x objective. Scale bars, 10 µm. (B-C) The perinuclear portion of the MK endoplasmic reticulum (ER) surrounds the internalized neutrophils (*s). The neutrophils are localized between the MK nucleus and ER. Note the inverse distribution of the DMS and ER. (D) Internalized neutrophils (*) did not colocalize with the Golgi apparatus of MKs.
Discussion
Emperipolesis is a cell-in-cell interaction at the interface of hemostasis and immunity. Neutrophils pass through MKs without disrupting the integrity of either cell, entering the MK cytoplasm in at least some cases. Other granulocytes, lymphocytes, erythrocytes, or monocytes are occasionally observed inside MKs, but neutrophils predominate and are observed more frequently than any other lineage, even after adjusting for their abundance in the bone marrow.24,40,41
Despite the ubiquity of EP across mammalian species, it remains unknown why neutrophils pass into MKs, whether different forms of EP exist, and what roles this interaction plays in health and disease. Understanding the cell biology of EP will be key to answering these questions. In the present study, we employed immunofluorescence and electron microscopy to study EP in a system that had previously been shown to model key elements of EP in vivo.24 Based on the duration of transit, we found that EP diverged into fast (generally <10 minutes) and slow (generally >60 minutes) forms. Neutrophils engaged in fast EP displayed ameboid motion, consistent with previous studies confirming a role for neutrophil cytoskeletal rearrangement in EP.24 By contrast, neutrophils engaged in slow EP assumed a rounded appearance, often near the megakaryocyte nucleus and surrounded by the perinuclear ER.
These observations strongly suggest that fast and slow EP are distinct, although without tools to block either form, specific physiologic roles remain difficult to define. Tavassoli et al26 suggested that bone marrow cells might take a transmegakaryocytic route to enter the circulation in states of increased cell demand. Indeed, an increase of EP has been observed in rodents after lipopolysaccharide-induced peritonitis, a condition of enhanced hematopoiesis requiring rapid mobilization of bone marrow cells.24,42 Fast EP could be suited for such a role, although the advantage of passing through an MK instead of directly through the thin sinusoid wall is unclear. Perisinusoidal MKs could simply offer more surface area for transit, complementing direct egress in a process analogous to how transendothelial migration complements egress between endothelial cells in peripheral tissues.43 Alternately, because MKs participate actively in EP, it may be that they select particular neutrophils for such passage, for example on the basis of surface integrin activation.24 An additional possibility is that EP allows MKs to modulate neutrophil function during passage. We had previously demonstrated that EP allows neutrophils and MKs to engage in bidirectional transfer of membranes between cytoplasmic neutrophils and the DMS, resulting in accelerated platelet production and the formation of “hybrid platelets” bearing neutrophil membrane.24 Although our EM studies again showed neutrophils directly within the MK cytoplasm, it remains possible that not all neutrophils take this course. Some neutrophils could transit through MKs while remaining within emperisomes and thus topologically outside the MK, akin to transendothelial migration.44 If so, opportunities for membrane exchange could be limited, although soluble material or exosomes might still transfer.
A role for MKs in egress to the circulation would be consistent also with our observation that EP is performed preferentially by the most mature MKs. During megakaryopoiesis, MK precursors differentiate into fully mature MKs in the vascular bone marrow niche.45 Depending on their maturity, MKs interact differently with endothelial cells. Although immature MKs mostly form smooth planar interfaces with endothelial cells, mature MKs protrude podosome-like structures and eventually cytoplasmic processes through the endothelial cell layer into the bone marrow sinusoids.46 Thus, preferential conduct of EP by mature MKs restricts the process to cells in direct contact with the lumen of blood vessels. Alternately, enhanced EP by mature MKs could reflect their larger cell size and more extensive DMS, providing a larger cell surface for cell-cell contact and more space to accommodate neutrophils.
Intriguingly, approximately half of neutrophils remain within MKs for an extended period, a process we term slow EP. These cells exhibit a sessile morphology. Failing to observe any degraded neutrophils within MKs, we assume that most eventually exit, though because almost all slow EP events extended beyond our video observations, we cannot confirm this assumption. De Pasquale et al35 proposed that EP might serve as a “sanctuary” for neutrophils in an unfavorable bone marrow environment, though why sanctuary should be necessary is unclear. Localization between the nucleus and the perinuclear ER raises additional novel possibilities, such as to “intercept” messenger RNA emerging from the nucleus or to modulate ER function.47,48 The identification of different types of emperipolesis also suggests different functions for this cell-in-cell interaction. Hence, emperipolesis observed at baseline (2% to 5% of MKs in healthy mice as determined by histology of thin sections24 ) could reflect “physiological” emperipolesis, whereas increased emperipolesis observed during myeloablative stress, hemorrhage, or inflammation21 could reflect “pathological” emperipolesis, with distinct cellular mechanisms and functions. Without direct evidence, all such possibilities remain purely speculative.
A central topological problem presented by EP is how neutrophils leave the emperisome to enter the MK cytoplasm.21,24 We approached this problem via EM of 45 EP events. The most common localization of neutrophils was within a clearly demarcated vesicle, termed the emperisome.24 In other instances, only a single membrane separated the cytoplasm of the neutrophil from that of the MK, consistent with intracytoplasmic residence (Figure 4Cii). We observed intermediate steps in which part of the neutrophil remained in the emperisome while part exhibited contact with the cell cytoplasm (Figure 4Ci). These images suggest penetration of the neutrophil through part of the vesicle wall rather than, for example, wholesale resorption or disintegration of the emperisome membrane. Other images are more difficult to categorize definitively with respect to emperisome vs cytoplasm, in particular where the neutrophil is surrounded by the DMS (Figure 4Bi-Bii). Earlier EM studies had described these cells as residing “loosely in the canalicular system,”49 though further study will be required to understand the topology with respect to the intravesicular, cytoplasmic, and extracellular compartments. The technical limitations of EM do not allow us to determine whether these morphological phases represent distinct neutrophil fates restricted to either fast or slow EP, or instead sequential stages undertaken by many or even all neutrophils during EP.
Our study has several important limitations. We studied murine EP, employing a useful but nevertheless in vitro system. How our findings translate to human and in vivo contexts remains unknown. Although fast and slow EP appear distinct from each other, suggesting distinct functions, we could not here define those functions and cannot exclude the possibility that they fulfill similar roles. Further research is required to elucidate how neutrophils transition between different intermediate stages, whether EP modulates the behavior of neutrophils, MKs, or platelets, and whether MKs outside the bone marrow compartment, for example in the lung, also engage neutrophils and other cells via EP.
Despite these limitations, our studies provide the first evidence that MK EP is a heterogeneous process through which neutrophils may engage with MKs either for a short or long duration, interacting with intracellular structures including the emperisome, the DMS, the cytoplasm, the perinuclear ER, and potentially the nucleus itself. Preserved in all mammalian species studied, across millions of years of otherwise divergent evolution,21 these observations suggest that EP will likely serve a range of roles to be defined through further investigations.
Acknowledgments
F.Y.H. was funded by a fellowship from Boehringer Ingelheim Fonds. P.C. was funded by the Arthritis National Research Foundation and the Gilead Sciences Research Scholars Program in Rheumatology. F.A.R. was funded by a fellowship from Boehringer Ingelheim Fonds. R.D. was funded by the Arthritis National Research Foundation. R.G.-B. was funded by a fellowship from Boehringer Ingelheim Fonds, a physician-scientist development grant from the Medical Faculty of Heidelberg, and a research grant from the German Society for Rheumatology. P.A.N. was supported by National Institutes of Health (NIH) National Institute of Arthritis and Musculoskeletal and Skin Diseases awards R01AR065538, R01AR075906, R01AR073201, R21AR076630, P30AR070253, and R56AR065538; NIH National Heart, Lung, and Blood Institute R21HL150575; the Fundación Bechara; and the Arbuckle Family Fund for Arthritis Research.
Authorship
Contribution: F.Y.H. designed and conducted experiments, analyzed data, and drafted the manuscript; P.C. designed, conducted, and supervised experiments, analyzed data, and edited the manuscript; F.A.R. conducted experiments and analyzed data; R.D. conducted experiments and analyzed data; R.G.-B. analyzed data and edited the manuscript; P.A.N. conceptualized and supervised the study, analyzed data, and drafted the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Peter A. Nigrovic, Division of Immunology, Boston Children’s Hospital, Karp Family Research Building, 10th Floor, One Blackfan Circle, Boston, MA 02115; e-mail: peter.nigrovic@childrens.harvard.edu.

