

Key Points
Fetal injection of antibodies against the c-Kit receptor and CD47 effectively depletes host HSCs in immunocompetent mice.
In utero depletion of host HSCs increases long-term engraftment after neonatal hematopoietic cell transplantation.
Introduction
In utero hematopoietic cell transplantation is a potential treatment for many genetic disorders, including hemoglobinopathies.1 Proposed benefits include transplanting into an immunologically naive fetal host with the ability to form donor-specific tolerance.2-4 Despite success in animal models,5-11 success in human clinical applications has been limited.12 Multiple barriers to engraftment have been elucidated, including the maternal immune system.8,13 Clinically, transplanting maternal cells may take advantage of existing fetal tolerance to maternal antigens and avoids the maternal immune barrier, a hypothesis that we are currently testing in a phase 1 clinical trial in fetuses with α thalassemia major.14 However, fetal transplantation does not involve preconditioning, and lack of space in the bone marrow niche is another potential barrier that must be overcome.15-17 We previously demonstrated that engraftment can be improved with in utero depletion of host hematopoietic stem cells (HSCs) by injecting monoclonal antibody against the c-Kit receptor (ACK2) into C57BL/6 mice.15 ACK2 blocks the interaction between the c-Kit receptor and stem cell factor ligand, resulting in a transient removal of endogenous HSCs, which depend on stem cell factor ligand for their survival,18 without depletion of mature immune cells. An ACK2-based antibody-depletion method is being used in a clinical trial for pediatric patients.19
Recently, simultaneous treatment with an anti-CD47 antibody was shown to potentiate the effect of ACK2 and result in much higher chimerism levels than seen in mice that received ACK2 alone.20 CD47 is expressed on HSCs and acts as a “don’t eat me” signal, preventing phagocytosis by macrophages and neutrophils via interaction with SIRPα.21,22 Blockade of the CD47–SIRPα interaction enhances antibody-dependent depletion, allowing the combination strategy to achieve depletion, even in wild-type mice. Here, we explore the safety and efficacy of this combination strategy in fetal mice.
Methods
Mice
Wild-type C57BL/6J (C57; CD45.2) and B6.SJL-PtrcaPep3b/BoyJ (BoyJ; CD45.1) mice were obtained from the National Cancer Institute. All mice were bred and maintained at the University of California, San Francisco. All procedures were performed according to the protocol approved by the University of California, San Francisco Institutional Animal Care and Use Committee.
In utero ACK2/MIAP410 injections
Pregnant dams were anesthetized at embryonic day 14.5 (E14.5) using isoflurane. A midline laparotomy was made, and the uterus was exteriorized. Fetal mice were injected with 2.5 μg of ACK2 or 2.5-μg concentrations of ACK2 and varying doses of MIAP410 (2.5 and 5 μg), or Dulbecco’s phosphate buffered saline (PBS) as controls, in a volume of 5 μL per fetus at E14.5. ACK2 antibody was purchased from BioLegend (San Diego, CA). MIAP410 was purchased from Bio X Cell (West Lebanon, NH).
Neonatal transplantation
Neonates were transplanted with congenic (B6.CD45.1/CD45.2) fetal liver mononuclear cells (FLMCs) on postnatal day 0 (P0) or P1. FLMCs were isolated from E13.5 or E14.5 donor fetuses by density gradient separation using Ficoll-Paque Plus (GE Healthcare), as previously described,3,8 and 2.5 × 106 FLMCs were resuspended in 10 µL of Dulbecco’s PBS and injected into the neonatal liver. Circulating chimerism levels were determined every 4 weeks by flow cytometry to enumerate CD45.1 (donor) and CD45.2 (host) cells.
Quantification of HSC depletion
Bone marrow mononuclear cells were harvested by dissection of neonatal femurs and tibias, and a single-cell suspension was made. The cells were then incubated with antibodies against lineage markers c-Kit and Sca-1 to quantify Lin−Sca-1+c-Kit+ (KLS) cells. Additional markers, including CD217, CD34, CD135, and CD16/32, were analyzed to identify progenitor populations. The same method was used for mice older than 24 weeks of age to confirm the presence of donor-derived HSCs.
Flow cytometric evaluation of myeloid cells
Spleens were harvested from mice at P1 after in utero preconditioning for analysis of erythroid development. A splenocyte single-cell suspension was created, and the cells were incubated with antibodies against CD45 and Gr-1.
Complete blood count
Whole blood was collected into EDTA capillary tubes (Fisher Scientific, Hampton, NH) via facial vein puncture in adult mice and at harvest in neonates. Samples were analyzed using a Hemavet 950 FS (Drew Scientific).
Tissue histology
Mice were harvested at 3 weeks of age, and liver, spleen, bone marrow, heart, kidney, intestine, and brain were collected. These underwent overnight fixation with 4% paraformaldehyde (Electron Microscopy Sciences), followed by overnight cryoprotection in 30% sucrose (Fisher Scientific). Tissue was then embedded in paraffin, sectioned at 4 μm, stained with hematoxylin and eosin, and reviewed by a pathologist.
Determination of chimerism levels
Blood was collected from the facial vein into heparinized tubes and washed, and red blood cells were lysed using ACK Lysing Buffer. Mononuclear cell preparations were incubated in fluorescence-activated cell-sorting staining buffer (PBS with 2% fetal bovine serum and 2 mM EDTA) with fluorochrome-conjugated antihuman surface antibodies. The following antibodies were used for flow cytometry: Pur-CD3 (17A2), Sca-1 (D7), CD71 (RI7217), Gr-1 (RB6-8C5), CD45.1 (A20), and CD3 (17A2) (BioLegend); Pur-CD8a (53-6.7), Pur-CD11b (M1/70), Pur-Ly6G (RB6-8C5), Pur-CD4 (GK1.5), Pur-CD45R (RA3-6B2), Pur-TER-119 (TER-119), Pur-CD5 (53-7.3), c-Kit (2B8), CD45.2 (104), and TER-119 (TER-119) (BioLegend); CD19 (1D3) (BD Pharmingen); and F(ab′)2 fragment of goat anti-rat IgG (H+L) and whole-molecule rat IgG (Invitrogen). Pur-CD117 (ACK2) and Pur-CD47 (MIAP410) were purchased from BioLegend and Bio X Cell, respectively. Cell viability was determined using a LIVE/DEAD Fixable Yellow Dead Cell Stain Kit (Molecular Probes). Flow cytometry was performed on an LSR II (BD) flow cytometer and analyzed using FACS Diva (BD) and FlowJo (TreeStar). Chimerism was expressed as the percentage of donor/(donor + host) cells. Mice with >1% circulating donor cells were considered chimeric. Lineage analysis of chimeric cells was performed by staining peripheral blood with antibodies against CD45.1, CD45.2, Gr-1, CD3, and CD19.
Statistics
Data comparing 2 groups were evaluated using the Student t test or the χ2 test. Multiple grouped data were evaluated using analysis of variance (ANOVA) with the Tukey Honest Significant Difference test, unless data were not normally distributed, in which case a Kruskal-Wallis test was performed. Data are shown as mean ± standard error of the mean. P < .05 was deemed statistically significant. Statistical analysis was performed using Prism GraphPad (GraphPad Software, La Jolla, CA).
Results and discussion
Fetal conditioning with c-Kit and CD47 blockade results in HSC depletion and improved engraftment
To determine the degree of host HSC depletion after ACK2 therapy or combination therapy with ACK2 and MIAP410, we harvested mice on P0, 7 days after in utero injection, and stained for KLS cells. ACK2 was kept at 2.5 μg, a dose that was effective with minimal toxicity in a previous study from our laboratory.15 However, the ACK2 used in this study was from a different manufacturer than the ACK2 evaluated in our previous study,15 and it was not effective in depleting host KLS cells when used alone. In contrast, combination therapy with ACK2 and MIAP410 showed significant decreases in total KLS cells at both doses of MIAP410 compared with ACK2 alone (Figure 1A-B). The addition of anti-CD47 improves KLS cell depletion by antibody-dependent cell-mediated phagocytosis, as described by Chhabra et al.20 Essentially, the effectiveness of ACK2 in depleting HSCs is dependent on cell-mediated toxicity from stem cell factor ligand blockade from the c-Kit receptor and antibody-mediated phagocytosis. CD47 is a marker of self that prevents phagocytosis by macrophages; when this “don’t eat me” signal is blocked with the MIAP410 antibody, cells become more susceptible to phagocytosis.
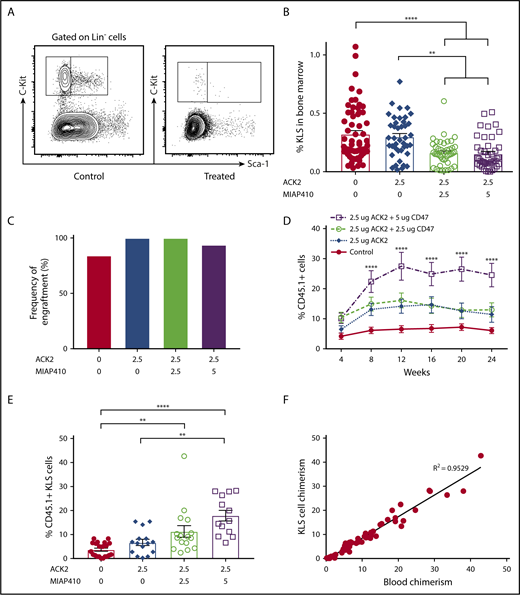
ACK2 and CD47 blockade results in KLS cell depletion in the bone marrow and improved engraftment of donor HSCs after congenic transplantation. (A) Representative gating strategy for identifying KLS cells. (B) Frequency of KLS cells within all live bone marrow cells harvested from bilateral femurs and tibias of P0 pups 7 days after treatment. (C) Rate of engraftment (number of animals with >1% chimerism/number of transplanted mice) at 4 weeks. PBS, n = 19; ACK2 (2.5 μg), n = 9; ACK2 (2.5 μg) + MIAP410 (2.5 μg), n = 18; ACK2 (2.5 μg) + MIAP410 (5 μg), n = 16. (D) Levels of donor CD45 chimerism in chimeric mice over the course of the study. The treatment group that received ACK2 (2.5 μg) + MIAP410 (5 μg) was found to have significantly higher chimerism compared with all other groups. The groups that received ACK2 alone and ACK2 + MIAP410 (2.5 μg) were found to be significantly more chimeric than PBS controls (P < .01 and P < .0001, respectively). PBS, n = 16; ACK2 (2.5 μg), n = 9; ACK2 (2.5 μg) + MIAP410 (2.5 μg), n = 18; ACK2 (2.5 μg) + MIAP410 (5 μg), n = 15. (E) Bone marrow chimerism of KLS cells at age ≥ 24 weeks. (F) Bone marrow KLS chimerism correlates with blood chimerism on linear regression. **P < .01, ****P < .0001, ANOVA with the Tukey multiple-comparison test.
ACK2 and CD47 blockade results in KLS cell depletion in the bone marrow and improved engraftment of donor HSCs after congenic transplantation. (A) Representative gating strategy for identifying KLS cells. (B) Frequency of KLS cells within all live bone marrow cells harvested from bilateral femurs and tibias of P0 pups 7 days after treatment. (C) Rate of engraftment (number of animals with >1% chimerism/number of transplanted mice) at 4 weeks. PBS, n = 19; ACK2 (2.5 μg), n = 9; ACK2 (2.5 μg) + MIAP410 (2.5 μg), n = 18; ACK2 (2.5 μg) + MIAP410 (5 μg), n = 16. (D) Levels of donor CD45 chimerism in chimeric mice over the course of the study. The treatment group that received ACK2 (2.5 μg) + MIAP410 (5 μg) was found to have significantly higher chimerism compared with all other groups. The groups that received ACK2 alone and ACK2 + MIAP410 (2.5 μg) were found to be significantly more chimeric than PBS controls (P < .01 and P < .0001, respectively). PBS, n = 16; ACK2 (2.5 μg), n = 9; ACK2 (2.5 μg) + MIAP410 (2.5 μg), n = 18; ACK2 (2.5 μg) + MIAP410 (5 μg), n = 15. (E) Bone marrow chimerism of KLS cells at age ≥ 24 weeks. (F) Bone marrow KLS chimerism correlates with blood chimerism on linear regression. **P < .01, ****P < .0001, ANOVA with the Tukey multiple-comparison test.
We next examined whether host HSC depletion with this strategy would improve engraftment. After in utero depletion with ACK2 or ACK2 + MIAP410 at various doses, we performed congenic fetal liver mononuclear cell transplantation (CD45.1/CD45.2 into CD45.2) of 2.5 million cells on P0 or P1. Blood chimerism levels were checked at 4-week intervals from age 4 weeks until harvest. Mice were considered engrafted if they had >1% donor cells; the percentage chimerism in engrafted animals was defined as donor CD45/(donor + host CD45) × 100%. Engraftment rates were similarly high among all groups (Figure 1C). Levels of chimerism were significantly higher in all depletion groups relative to PBS controls (Figure 1D). Interestingly, although ACK2 alone did not demonstrate depletion of the KLS cells, engraftment levels were also higher in the ACK2 group compared with controls, suggesting that our use of KLS cell numbers (which was chosen over enumerating CD48+/CD150+ cells because of the low cell numbers in neonatal bone marrow) as a surrogate for bone marrow stem cells was not sensitive enough to detect subtle differences in depletion. The addition of 2.5 μg of MIAP to ACK2 did not result in higher levels of engraftment. However, fetal conditioning with ACK2 + 5 μg of MIAP resulted in significantly higher peripheral blood chimerism compared with mice treated with PBS, ACK2 alone, or ACK2 + 2.5 μg of MIAP410.
Given the stable levels of chimerism over time, we next examined KLS cell engraftment in the bone marrow at or after week 24. The bone marrow KLS chimerism levels mirrored the stepwise increase in blood chimerism (Figure 1E). Moreover, the levels of KLS cells in the bone marrow correlated closely with the blood chimerism levels (Figure 1F). We also examined other donor-derived progenitor populations in the bone marrow of adult mice and found improved chimerism with combination ACK2 and MIAP410 therapy relative to PBS controls in all progenitor populations examined, including long-term HSCs, short-term HSCs, multipotent progenitors, and granulocyte-macrophage progenitors (supplemental Figure 1). Although we did not specifically evaluate erythroid chimerism in this congenic transplant method, we evaluated the common myeloid progenitor cells and megakaryocyte erythrocyte progenitors that eventually give rise to erythrocytes and found improved chimerism among them as well (supplemental Figure 1).
Toxicity of fetal conditioning with ACK2 and MIAP410
We evaluated the overall survival to birth of pups after ACK2 treatment and combination therapy with ACK2 and MIAP410. We found that, although ACK2 alone did not decrease survival, the addition of MIAP410 (5 μg) did (Figure 2A). This decreased survival was associated with anemia in surviving pups (Figure 2B). These results are consistent with previous reports showing a decrease in circulating erythrocytes after CD47 blockade.23 Analysis of pups receiving the higher dose of MIAP410 (5 μg) revealed significant anemia 4 days after in utero injection (prior to their demise, on E18, Figure 2C), indicating that anemia is the likely reason for the low survival seen in this group. Histologic analysis of various tissues in surviving mice (ACK2 [2.5 μg] + MIAP410 [5 μg]; n = 12, PBS n = 18) did not show any toxicity to liver, spleen, kidney, heart, lung, bone marrow, intestines, or brain. In the treatment and control groups, we did note brown pigmented spindled cells in the spleen, negative for hemosiderin on Prussian Blue staining, which likely reflects physiologic splenic melanosis that is seen intermittently in C57BL/6 mice.24
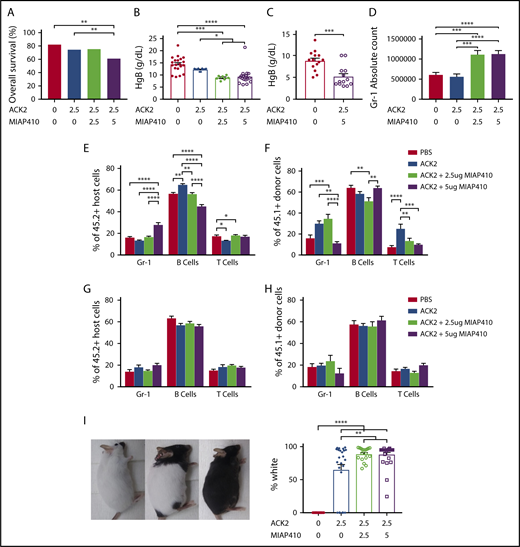
In utero blockade of c-Kit and CD47 increases Gr-1+cells and prevents early engraftment of Gr-1 cells, in addition to causing transient anemia and permanent fur whitening. (A) Survival to birth after in utero injection of PBS, ACK2, or ACK2 + CD47 at various concentrations. PBS, n = 128; ACK2 (2.5 μg), n = 94; ACK2 (2.5 μg) + MIAP410 (2.5 μg), n = 122; ACK2 (2.5 μg) + MIAP410 (5 μg), n = 154. **P < .01, χ2 test. Hemoglobin levels in blood of P0 mice (B) and E18 mice (C) after in utero HSC depletion. (D) Numbers of Gr-1+ cells in the spleens of P0 mice. Lineage distribution of host-derived (E) and donor-derived (F) cells in blood from 4-week-old mice. Lineage distribution of host-derived (G) and donor-derived (H) cells in blood of 12-week-old mice. (I) Hypopigmentation in mice after undergoing in utero HSC depletion. In panels B-F,I, *P < .05, **P < .01, ***P < .001, ****P < .0001, ANOVA with the Tukey multiple-comparison test.
In utero blockade of c-Kit and CD47 increases Gr-1+cells and prevents early engraftment of Gr-1 cells, in addition to causing transient anemia and permanent fur whitening. (A) Survival to birth after in utero injection of PBS, ACK2, or ACK2 + CD47 at various concentrations. PBS, n = 128; ACK2 (2.5 μg), n = 94; ACK2 (2.5 μg) + MIAP410 (2.5 μg), n = 122; ACK2 (2.5 μg) + MIAP410 (5 μg), n = 154. **P < .01, χ2 test. Hemoglobin levels in blood of P0 mice (B) and E18 mice (C) after in utero HSC depletion. (D) Numbers of Gr-1+ cells in the spleens of P0 mice. Lineage distribution of host-derived (E) and donor-derived (F) cells in blood from 4-week-old mice. Lineage distribution of host-derived (G) and donor-derived (H) cells in blood of 12-week-old mice. (I) Hypopigmentation in mice after undergoing in utero HSC depletion. In panels B-F,I, *P < .05, **P < .01, ***P < .001, ****P < .0001, ANOVA with the Tukey multiple-comparison test.
We also found that in utero ablation resulted in inflammation in the fetal host, with increased host Gr-1+ cells in the spleens at 1 week after injection in mice that received combination therapy with ACK2 and MIAP410 (Figure 2D). The increase in host Gr-1+ cells persisted at 4 weeks (Figure 2E), resulting in comparatively lower engraftment of donor Gr-1+ cells at this time point (Figure 2F). However, these changes were transient, and the lineage distribution of engrafted cells was normalized among all groups by 12 weeks (Figure 2G-H).
Finally, we demonstrated that most of the mice that received fetal conditioning developed severe coat discoloration (Figure 2I). In our previous study, using a different source of ACK2, we had seen altered coat color when ACK2 was given on E13.5, but not when given on E14.5,15 and only in a minority of the treated animals. Treatment with ACK2 affects the multiple stages of melanocyte development, likely due to its effect on c-Kit receptors found on melanocytes during fetal melanocyte migration.25-27 Melanocyte migration to the dermis requires interaction with stem cell factor and c-Kit receptor.28 Interestingly, a small percentage of the mice that received ACK2 alone did not have any coat discoloration (Figure 2I), whereas whitened fur was seen uniformly when MIAP410 was added. We speculate that MIAP410 potentiates the effect of ACK2 by enhancing phagocytosis of melanocytes, similar to its effects on HSC depletion.
Our study demonstrates that antibody-mediated depletion of host HSCs with c-Kit and CD47 blockade improves systemic engraftment of congenic HSCs after in utero transplantation without the need for host irradiation. We also demonstrate that the systemic engraftment is long term and present in multiple hematopoietic progenitor populations. Because there is currently a clinical trial of c-Kit blockade in pediatric patients with severe combined immunodeficiency19 and a clinical trial of in utero transplantation of maternal HSCs in fetuses with α thalassemia major,14 our results have clinical implications. Fetal conditioning prior to in utero transplantation may be a viable strategy to improve engraftment, particularly in fetuses with α thalassemia major that are already receiving multiple blood transfusions to combat treatment-related anemia. Fetal conditioning would precede transplantation of maternal stem cells by 1 to 2 weeks, if the half-life of the ACK2 antibody is determined to be similar in large animal models of fetal therapy. However, the pervasiveness of c-Kit expression increases the risk of off-target effects, particularly in melanocytes. Because the degree of melanocyte toxicity was dependent on the lot of antibody used, similar assays will need to be performed with the clinical-grade antibody in large animal models to determine the safety of this strategy.
The full-text version of this article contains a data supplement.
Acknowledgments
The authors thank Agnieszka Czechowicz and members of the MacKenzie laboratory for helpful discussions.
This work was supported by an Eli and Edythe Broad Stem Cell Fellowship (R.G.W.) and by a California Institute of Regenerative Medicine New Physician Scientist Translational Research Award (RN3-06532).
Authorship
Contribution: T.C.M. and R.G.W. designed the experiments; R.G.W., Q.-H.N., C.E., and B.W. .performed experiments and analyzed data; A.N.M. performed histologic tissue analysis; and R.G.W., Q.-H.N., and T.C.M. wrote the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Tippi C. MacKenzie, Department of Surgery, University of California, San Francisco, 550 16th St, San Francisco, CA 94143-0570; e-mail: tippi.mackenzie@ucsf.edu.

