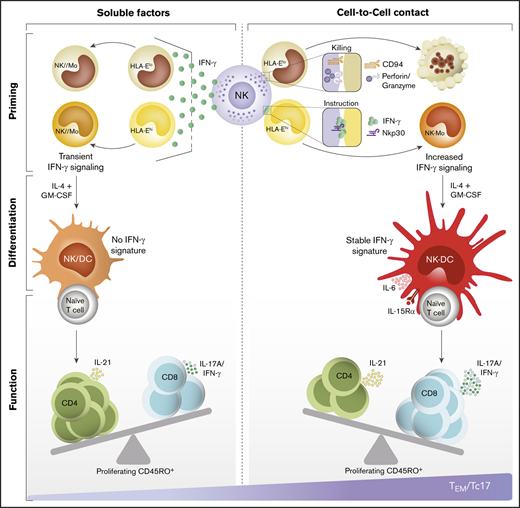
Key Points
Early interactions between NK cells and CD14+CD16− monocytes determine the functional specialization of subsequent monocyte-derived DCs.
NK-cell–trained DCs drive the priming of Tc17 responses, which are decreased in blood from patients with classical NK-cell deficiency.

Abstract
Adaptive immune responses are acknowledged to evolve from innate immunity. However, limited information exists regarding whether encounters between innate cells direct the generation of specialized T-cell subsets. We aim to understand how natural killer (NK) cells modulate cell-mediated immunity in humans. We found that human CD14+CD16− monocytes that differentiate into inflammatory dendritic cells (DCs) are shaped at the early stages of differentiation by cell-to-cell interactions with NK cells. Although a fraction of monocytes is eliminated by NK-cell–mediated cytotoxicity, the polarization of interferon-γ (IFN-γ) at the NKp30-stabilized synapses triggers a stable IFN-γ signature in surviving monocytes that persists after their differentiation into DCs. Notably, NK-cell–instructed DCs drive the priming of type 17 CD8+ T cells (Tc17) with the capacity to produce IFN-γ and interleukin-17A. Compared with healthy donors, this cellular network is impaired in patients with classical NK-cell deficiency driven by mutations in the GATA2 gene. Our findings reveal a previously unrecognized connection by which Tc17-mediated immunity might be regulated by NK-cell–mediated tuning of antigen-presenting cells.
Introduction
In early inflammation, the rapid activation of natural killer (NK) cells influences adjacent innate cells, impacting the course of immunity. Endowed with a complex repertoire of activating and inhibitory receptors, NK cells exert a variety of functions1 ; although they were discovered by their ability to kill tumor and virus-infected cells in the absence of previous stimuli,2,3 they also play significant roles in immune tolerance and in the regulation of other cells through the production of cytokines and chemokines.4,5 A well-known example is their cooperative cross talk with dendritic cells (DCs), which precedes the activation and expansion of specialized T cells.6,7 This dialogue promoted by the coexistence of both cells in inflamed tissues is initiated by the recognition of activating ligands by either NK cells or DCs.8-10 Through the production of interleukin-12 (IL-12), interferon-α/β (IFN-α/β), IL-15, and IL-18, DCs promote survival, proliferation, and activation of NK cells, which in turn, increase their cytotoxic capacity and ability to produce cytokines.6,11-13 Depending on the timing and intensity of this cross talk, NK cells can tune cell-mediated immunity by either activating or killing immature DCs via the NKp30 receptor.6-8,14 Through the production of tumor necrosis factor (TNF) and IFN-γ, activated NK cells induce the maturation of DCs, characterized by an exceptional ability to produce IL-12 and to promote T helper type 1 (Th1) and cytotoxic T lymphocyte (CTL) responses.6,7,15,16 As a mechanism to either amplify or restrain DC-mediated inflammation, poorly immunogenic immature DCs are targets of NK-cell–mediated cytotoxicity, thus selecting highly immunogenic DCs and constraining DC-mediated immune responses by the elimination of immature precursors.6,8,17
Studies in mice have shown that NK cells also regulate monocytes. The timely production of type I interferon after mucosal infection by HSV-2 induces CCL2-mediated recruitment of IL-18–producing inflammatory monocytes, essential for NK-cell–mediated IFN-γ production and viral control.18 During Toxoplasma gondii infection, the IFN-γ secreted by NK cells allows the replacement of resident phagocytes by circulating monocytes that differentiate in situ into IL-12–producing DCs, required to initiate the response against the pathogen.19 During T gondii oral infection, the systemic effect of IL-12 activates bone marrow–resident NK cells to produce IFN-γ, which promotes the priming of monocyte precursors with regulatory function. These regulatory monocytes when released to the blood stream reach the infection site and promote inflammation resolution through IL-10 production.20
In humans, quantitative proteomics-based analyses of blood cells at steady and activated states provided insight on such complex networks of intercellular communication and suggested previously unrecognized connections between NK cells and the orchestration of immunity.21 For instance, the presence of the NK-cell–DC axis in human tumors correlates with enhanced T-cell responses, patient responsiveness to checkpoint inhibitor immunotherapy, and overall survival.22 Yet, how NK cells are linked to specialized T-cell responses remains unclear. Here, we describe how human NK cells regulate early stages of monocyte differentiation and subsequently impact the ability of monocyte-derived DCs (Mo-DCs) to drive T-cell specialization.
Methods
Study approval
This study was approved by the National Institutes of Health (NIH) Clinical Center Department of Transfusion Medicine (99-CC-0168), the Institutional Review Board of Baylor College of Medicine (H-30487), and the Institutional Review Board of Hospital de Clinicas de Porto Alegre (2010-0457). All samples were obtained after patients’ deidentification and signed written informed consent.
Cell isolation
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats from male healthy donors (HDs), aged between 19 and 45 years, after centrifugation gradient with Ficoll-Paque PLUS (GE Healthcare). Untouched NK cells were obtained using the Human NK-cell isolation kit (Miltenyi Biotec). For activation, NK cells were treated for 6 hours with 10 ng/mL IL-15 (PeproTech) and 250 IU/mL IL-2 (Hoffmann-La-Roche, Inc). Untouched CD16−CD14+ monocytes were isolated using the Human Monocyte Isolation kit II (Miltenyi Biotec). Purity obtained ranged between 96% and 99%; remaining platelets accompanied all monocyte fractions.
Monocytes and NK-cell cocultures
Monocytes were cultured alone (Mo) or cocultured with preactivated NK cells in V-bottom plates at a 5:1 ratio for 18 hours, having direct contact (NK-Mo) or separated through 96-Transwell membranes (0.4-μm pore-size; Corning) (NK//Mo). Monocytes cocultured directly were depleted of NK cells by magnetic separation using the Human CD56 Microbeads kit (Miltenyi Biotec) before subsequent differentiation. Some experiments were performed in the presence of blocking antibodies against NKp30 (P30-15), IFN-γ (MD-1), CD94 (DX22), or immunoglobulin G1,κ (IgG1,κ) isotype control (all LEAF from BioLegend). In some experiments, monocytes were pretreated with 1000 IU/mL IFN-γ (PeproTech) for 18 hours.
Confocal microscopy
Deep red (Life Technologies) prestained monocytes and preactivated NK cells were cocultured in medium containing 0.1% Live/Dead blue (Life Technologies), for 6 hours in a µ-Slide 8 Well Glass Bottom (Ibidi). Cells were fixed, permeabilized, and stained with AF488–IFN-γ (D3H2; Cell Signaling) and AF546-Phalloidin (Invitrogen). Images were acquired using a Zeiss LSM 880/Airyscan, rendered, and analyzed using IMARIS (8.2.0) and Zen2012 (Zeiss). For live-cell imaging, cocultures of monocytes (deep red) and NK cells (carboxyfluorescein diacetate succinimidyl ester [CFSE]) were incubated in the XL-3 incubator (PeCon GmbH); interactions recorded every 20 minutes for 12 hours by the LSM-510-NLO (Zeiss) and analyzed using Zen2009 (Zeiss).
Mo-DCs and T-cell cocultures
Monocytes were differentiated into DCs using 50 ng/mL each of IL-4 and granulocyte-macrophage colony-stimulating factor (GM-CSF; PeproTech) for 5 days; DCs were activated with 50 ng/mL TNF-α (PeproTech) for 24 hours. At harvest, Mo-DCs were cocultured for 5 days, 1:10 with allogeneic naive T cells prepurified using the Naive-Pan-T-cell isolation kit (Miltenyi Biotec). In some experiments, 10 ng/mL IL-6 and 10 ng/mL IL-15 (PeproTech) were added to cultures.
Flow cytometry
Phenotypic and functional changes in NK cells, monocytes, Mo-DCs, and T cells were monitored by flow cytometry (supplemental Materials and methods) using the BD LSRFortessa (BD Biosciences) and analyzed using FlowJo (FlowJo LLC). Monocytes and DCs were sorted using the MoFlo Astrios (Beckman Coulter, Inc) before gene-expression analysis.
RNA extraction and gene-expression analysis
Total RNA from presorted monocytes and Mo-DCs were hybridized to the Human GeneChip 2.0 ST array (Affymetrix) and analyzed using different tools (supplemental Materials and methods). Gene expression was validated by real-time polymerase chain reaction (PCR) using the QuantStudio-6-Flex and analyzed using the QuantStudioTM Real-Time PCR software 1.1 (Applied Biosystems).
Single-cell RNA sequencing
PBMC libraries derived from GATA2-deficient patients and HD controls were prepared using Chromium Single Cell 5′ Library Construction kit (10X Genomics). Single-cell RNA sequencing (scRNA-seq) libraries were pooled and sequenced using the NextSeq 500/550 kit and NextSeq system (Illumina). Sequencing reads were processed and aggregated using Cell Ranger (10X Genomics) and data were analyzed using Partek Flow (Partek) and SeqGeq (FlowJo).
Statistics
Data were analyzed using GraphPad Prism software 5.0 (GraphPad Software). Differences between groups were estimated using the unpaired Student t test with Welch correction. Values of P ≤ .05 were considered statistically significant.
Results
Cell-to-cell contact with NK cells triggers the IFN-γ–signaling pathway in CD14+CD16− monocytes
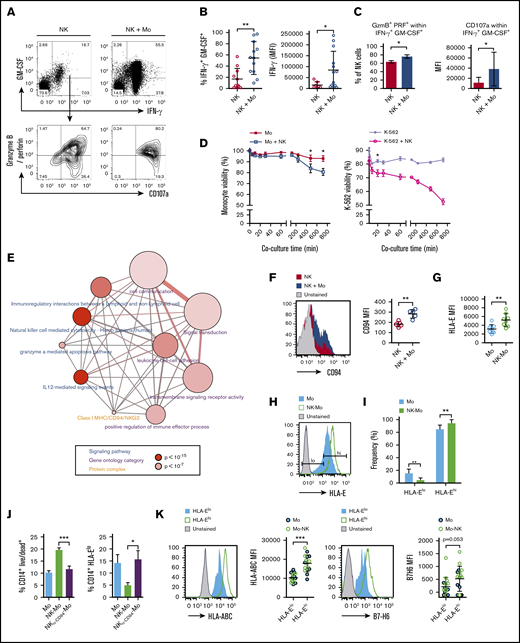
During inflammation, common chemokine networks rapidly and simultaneously recruit blood-borne NK cells and monocytes to tissues.23,24 We reproduced this encounter in vitro and assessed its impact in the transcriptional, phenotypic, and functional profile of classical CD14+CD16− monocytes. Monocytes were cocultured for 18 hours with preactivated NK cells, allowing cell-to-cell contact (NK-Mo) or keeping them separated by a Transwell membrane (NK//Mo). The IFN-γ–signaling pathway was upregulated in monocytes after contact with NK cells (Figure 1A; supplemental Figure 1). Accumulation of STAT1 and IRF1 gene transcripts increased in monocytes directly exposed to cell-to-cell contact with NK cells and, at a lower level, in those separated by NK cells through a Transwell membrane (Figure 1B; supplemental Figure 1). IFN-γ–inducible CXCL10 and CXCL9 gene transcription only increased upon cell-to-cell contact (Figure 1C). Similarly, transcript accumulation and protein expression of the IFN-γ–inducible CD64 (FCGRA1)25 increased only after cell-to-cell contact (Figure 1D), as CX3CR126 (Figure 1E), which was prevented by neutralization of IFN-γ (Figure 1F). Lipopolysaccharide (LPS) challenge showed that monocyte capacity to produce TNF and IL-23 was preconditioned by direct contact with NK cells, but in contrast to TNF, blocking IFN-γ did not significantly impact IL-23 production (Figure 1G).
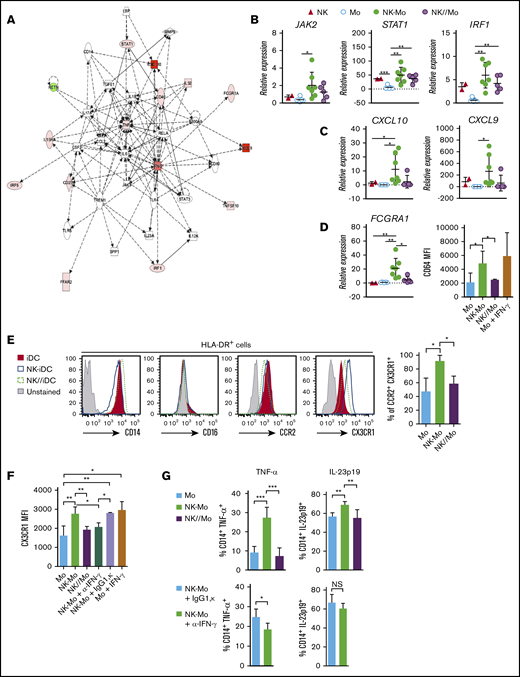
CD14+CD16−monocytes acquire stable IFN-γ signature only after cell-cell contact with NK cells. CD14+CD16− monocytes and in vitro preactivated NK cells were isolated from HD blood and cocultured for 18 hours (NK-Mo). Monocytes alone (Mo) or cocultured with NK cells through a Transwell membrane (NK//Mo) were used as controls. In some experiments, controls included monocytes treated with IFN-γ (Mo + IFN-γ). Some coculture experiments were done in the presence of anti-IFN-γ (NK-Mo + α-IFN-γ) or IgG1,κ isotype control (NK-Mo + IgG1,κ). After coculture, the transcriptional, phenotypical, and functional profiles of monocytes were assessed. (A) Main gene signatures induced in monocytes after coculture with NK cells, assessed by microarray. (B-C) Validation by real-time PCR of JAK2, STAT1, IRF1, CXCL10, and CXCL9 gene expression in monocytes after coculture with NK cells. (D) Validation of FCGRA1 gene expression and CD64 protein expression in monocytes after coculture with NK cells assessed by real-time PCR and flow cytometry, respectively. (E) Histograms for phenotypical characterization of monocytes after coculture with NK cells; frequency of CCR2+CX3CR1+ monocytes assessed by flow cytometry. (F) Expression (MFI) of CX3CR1 in monocytes after coculture with NK cells, assessed by flow cytometry. (G) Frequency of TNF-α+ and IL-23p19+ monocytes after LPS stimulation, assessed by flow cytometry. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001. NS, not significant.
CD14+CD16−monocytes acquire stable IFN-γ signature only after cell-cell contact with NK cells. CD14+CD16− monocytes and in vitro preactivated NK cells were isolated from HD blood and cocultured for 18 hours (NK-Mo). Monocytes alone (Mo) or cocultured with NK cells through a Transwell membrane (NK//Mo) were used as controls. In some experiments, controls included monocytes treated with IFN-γ (Mo + IFN-γ). Some coculture experiments were done in the presence of anti-IFN-γ (NK-Mo + α-IFN-γ) or IgG1,κ isotype control (NK-Mo + IgG1,κ). After coculture, the transcriptional, phenotypical, and functional profiles of monocytes were assessed. (A) Main gene signatures induced in monocytes after coculture with NK cells, assessed by microarray. (B-C) Validation by real-time PCR of JAK2, STAT1, IRF1, CXCL10, and CXCL9 gene expression in monocytes after coculture with NK cells. (D) Validation of FCGRA1 gene expression and CD64 protein expression in monocytes after coculture with NK cells assessed by real-time PCR and flow cytometry, respectively. (E) Histograms for phenotypical characterization of monocytes after coculture with NK cells; frequency of CCR2+CX3CR1+ monocytes assessed by flow cytometry. (F) Expression (MFI) of CX3CR1 in monocytes after coculture with NK cells, assessed by flow cytometry. (G) Frequency of TNF-α+ and IL-23p19+ monocytes after LPS stimulation, assessed by flow cytometry. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001. NS, not significant.
The CD14+CD16− monocyte compartment is shaped by NK-cell–mediated cytotoxicity
Following coculture with monocytes, the frequency of NK cells producing IFN-γ and GM-CSF increased (Figure 2A-B), along with granzyme B and perforin, concomitantly upregulating the degranulation marker CD107a (Figure 2C). Monocyte viability decreased ∼10% after 6 hours of coculture with NK cells, whereas similar degree of cell death was observed in K-562 cell after 20 minutes of coculture. (Figure 2D). Transcriptome analysis revealed NK-cell–related genes in the NK-primed monocytes fraction (supplemental Table 1), confirmed by real-time PCR (supplemental Figure 1; ie, IFNG gene); because NK-cell contamination of sorted monocytes was <1%, we hypothesized this was due to messenger RNA (mRNA) exchange from NK cells to monocytes, such as mRNA-loaded exosomes or mRNA trafficking via membrane nanotubes.27,28 Using an overrepresentation pathway-based analysis,29 we predicted from upregulated NK-related genes (supplemental Table 1) that regulatory and cytotoxic interactions occurred within the cells, initiated by IL-12 and dependent on cell-to-cell adhesion (Figure 2E). The analysis revealed a protein-protein complex of major histocompatibility complex (MHC) class I molecules and the CD94/NKG2 family, particularly, high expression of NKG2E gene transcripts (supplemental Table 1). The activating CD94/NKG2E receptor binds nonclassical MCH class I molecule, HLA-E, which is also recognized by the inhibitory receptor CD94/NKG2A.30 Low expression of HLA-E leads to target lysis31 whereas high levels protect targets from NK-cell cytotoxicity.32 CD94 molecules were upregulated in monocyte-exposed NK cells (Figure 2F) whereas HLA-E molecules were upregulated in NK-primed monocytes (Figure 2G). In controls, we observed a population of HLA-Elow monocytes that disappeared in NK-primed monocytes (Figure 2H-I). CD94 blockage during coculture prevented both NK-cell–mediated monocyte-death and loss of HLA-E low-expressing monocytes (Figure 2J). Regardless of the group (Mo or Mo-NK), total HLA-Elow monocytes express low levels of HLA-ABC and B7-H6, the NKp30 ligand33 (Figure 2K), suggesting that additional NK receptors could be involved in the killing and licensing of monocytes.

CD14+CD16−monocyte compartment is shaped by NK-cell–mediated cytotoxicity. CD14+CD16− monocytes and preactivated NK cells were isolated from HD blood and cocultured for 18 hours (NK-Mo). After coculture, NK cells’ capacity to produce cytotoxic proteins and cytokines after PMA plus ionomycin stimuli was assessed by flow cytometry. NK cells alone were used as controls. (A) Gating strategy for the analysis of GM-CSF+IFN-γ+ and granzyme B/perforin+CD107a+ NK cells. (B) Frequency of GM-CSF+ IFN-γ+ NK cells; production (iMFI: frequency multiplied by MFI) of IFN-γ by NK cells after contact with monocytes. (C) Frequency of granzyme B/perforin+ within GM-CSF+IFN-γ+ NK cells; expression (MFI) of CD107a by NK cells after contact with monocytes. Cytotoxic function of NK cells was monitored during the cocultures through the assessment of target viability by flow cytometry. K-652 cells were used as positive controls for NK-cell–induced cytotoxicity. (D) Monocyte viability during coculture (5-720 minutes) with NK cells; K-562 viability during the coculture (5-720 minutes) with NK cells. (E) Overrepresentation pathway-based analysis29 of NK-cell–related genes found upregulated in the transcriptome of NK-cell–primed monocytes. The input gene list (supplemental Table 1) was compared with 13-pathway databases considering a minimum overlap of 2 genes and P < .01. Each node represents a different pathway (blue), ontology category (purple), or protein-protein complex (orange); node size represents the number of genes contained in the set of a particular pathway and node color represents the P value. Nodes connected through edges share members and edges width reflect overlapping of genes between nodes. (F) Expression (MFI) of CD94 in NK cells after coculture with monocytes. (G) Expression (MFI) of HLA-E in monocytes after coculture with NK cells. (H) Gating strategy for the analysis of monocytes expressing low and high levels of HLA-E. (I) Frequency of HLA-Elow and HLA-Ehigh monocytes after coculture with NK cells. Monocyte viability was assessed by flow cytometry after contact with NK cells; for some groups, the CD94 receptor was blocked prior the interaction with monocytes (NKα-CD94-Mo). (J) Frequency of CD14+ monocytes positive for Live/Dead dye; frequency of CD14+HLA-Elow monocytes. (K) Histograms and pooled expression (MFI) of HLA-ABC and B7-H6 in HLA-Elow and HLA-Ehigh in monocytes from both groups (Mo and NK-Mo). Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001.
CD14+CD16−monocyte compartment is shaped by NK-cell–mediated cytotoxicity. CD14+CD16− monocytes and preactivated NK cells were isolated from HD blood and cocultured for 18 hours (NK-Mo). After coculture, NK cells’ capacity to produce cytotoxic proteins and cytokines after PMA plus ionomycin stimuli was assessed by flow cytometry. NK cells alone were used as controls. (A) Gating strategy for the analysis of GM-CSF+IFN-γ+ and granzyme B/perforin+CD107a+ NK cells. (B) Frequency of GM-CSF+ IFN-γ+ NK cells; production (iMFI: frequency multiplied by MFI) of IFN-γ by NK cells after contact with monocytes. (C) Frequency of granzyme B/perforin+ within GM-CSF+IFN-γ+ NK cells; expression (MFI) of CD107a by NK cells after contact with monocytes. Cytotoxic function of NK cells was monitored during the cocultures through the assessment of target viability by flow cytometry. K-652 cells were used as positive controls for NK-cell–induced cytotoxicity. (D) Monocyte viability during coculture (5-720 minutes) with NK cells; K-562 viability during the coculture (5-720 minutes) with NK cells. (E) Overrepresentation pathway-based analysis29 of NK-cell–related genes found upregulated in the transcriptome of NK-cell–primed monocytes. The input gene list (supplemental Table 1) was compared with 13-pathway databases considering a minimum overlap of 2 genes and P < .01. Each node represents a different pathway (blue), ontology category (purple), or protein-protein complex (orange); node size represents the number of genes contained in the set of a particular pathway and node color represents the P value. Nodes connected through edges share members and edges width reflect overlapping of genes between nodes. (F) Expression (MFI) of CD94 in NK cells after coculture with monocytes. (G) Expression (MFI) of HLA-E in monocytes after coculture with NK cells. (H) Gating strategy for the analysis of monocytes expressing low and high levels of HLA-E. (I) Frequency of HLA-Elow and HLA-Ehigh monocytes after coculture with NK cells. Monocyte viability was assessed by flow cytometry after contact with NK cells; for some groups, the CD94 receptor was blocked prior the interaction with monocytes (NKα-CD94-Mo). (J) Frequency of CD14+ monocytes positive for Live/Dead dye; frequency of CD14+HLA-Elow monocytes. (K) Histograms and pooled expression (MFI) of HLA-ABC and B7-H6 in HLA-Elow and HLA-Ehigh in monocytes from both groups (Mo and NK-Mo). Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001.
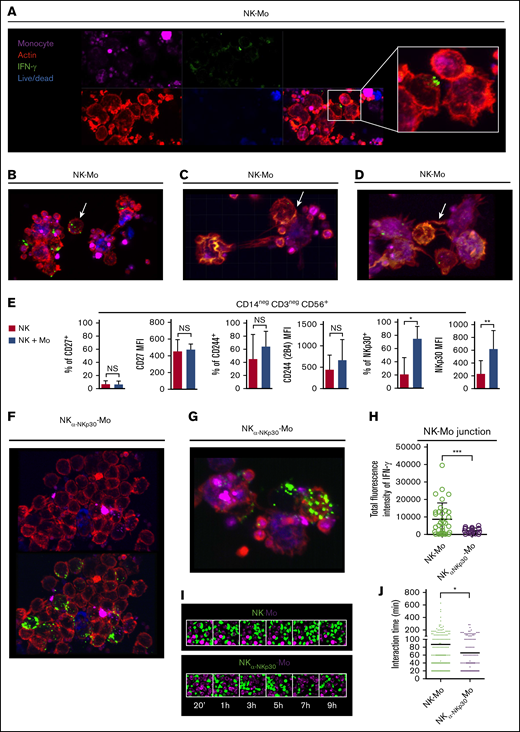
Surviving CD14+CD16− monocytes are instructed by IFN-γ polarization at the NKp30-stabilized synapses with NK cells
The elimination of monocytes expressing low levels of MHC class I molecules by NK-cell–mediated cytotoxicity may explain the role of contact to shape a monocyte population with distinct features; however, it does not explain why surviving monocytes rely on contact with NK cells to acquire an IFN-γ signature. To investigate this, we imaged the IFN-γ production during interaction of monocytes with NK cells. We observed that when the cells are in close contact, IFN-γ clustered in the membrane of NK cells toward monocytes and high content of IFN-γ was present within cell-to-cell junctions (Figure 3A). This polarization only occurs when NK cells are contiguous to monocytes because no polarization is observed in nonagglomerated NK cells (Figure 3B). IFN-γ clusters are also found within the junction of NK cells and dying monocytes (Figure 3C). We also observed nanotube-like structures that may be involved in transporting molecules between NK cells and monocytes possibly including mRNA or IFN-γ (Figure 3D; supplemental Figure 2A).
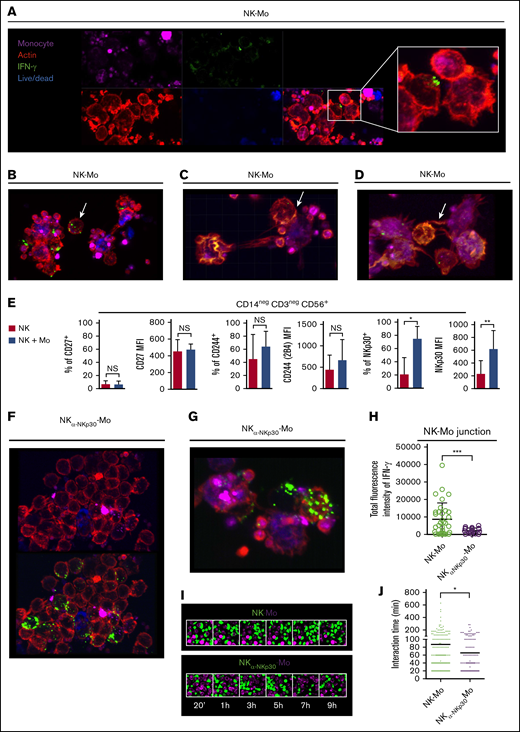
CD14+CD16−monocytes are instructed by the polarization of IFN-γ at the NKp30-mediated junction with NK cells. Preactivated NK cells and CD14+CD16− prestained monocytes (violet), isolated from HD blood, were cocultured for 6 hours in the presence of Live/Dead viability dye (blue). After coculture, cells were fixed and stained for IFN-γ (green) and phalloidin (red). In all experiments and groups, remaining platelets accompanied the monocyte fractions. For some cocultures, the NKp30 receptor was blocked prior the interaction with monocytes (NKα-NKp30-Mo). (A) IFN-γ dynamics during the interaction of NK cells with monocytes. (B) Distribution of IFN-γ in nonagglomerated NK cells (arrow). (C) Distribution of IFN-γ during the interaction of NK cells with dying monocytes (arrow). (D) Presence of IFN-γ in nanotube-like structures (arrow). (E) Frequency and expression (MFI) of CD27, CD244 and NKp30 on NK cells cultured alone (NK) or after 18 hours of coculture with CD14+CD16− monocytes (NK + Mo). (F) Distribution of IFN-γ during the interaction of NKp30-blocked NK cells with monocytes. (G) IFN-γ dynamics during the interaction of NKp30-blocked NK cells with monocytes. (H) Total fluorescence intensity of IFN-γ determined at the junction formed between NK cells and monocytes. Prestained NK cells (green) and monocytes (violet) were cocultured and live-cell imaging was recorded for up to 12 hours. (I) Captures of the interaction between NK cells and monocytes. (J) Interaction time of monocytes with NK cells vs monocytes with NKp30-blocked NK cells. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001. See “Confocal microscopy” for stains used; original magnifications ×20 (A-B,F,I) and ×40 (C-D,G).
CD14+CD16−monocytes are instructed by the polarization of IFN-γ at the NKp30-mediated junction with NK cells. Preactivated NK cells and CD14+CD16− prestained monocytes (violet), isolated from HD blood, were cocultured for 6 hours in the presence of Live/Dead viability dye (blue). After coculture, cells were fixed and stained for IFN-γ (green) and phalloidin (red). In all experiments and groups, remaining platelets accompanied the monocyte fractions. For some cocultures, the NKp30 receptor was blocked prior the interaction with monocytes (NKα-NKp30-Mo). (A) IFN-γ dynamics during the interaction of NK cells with monocytes. (B) Distribution of IFN-γ in nonagglomerated NK cells (arrow). (C) Distribution of IFN-γ during the interaction of NK cells with dying monocytes (arrow). (D) Presence of IFN-γ in nanotube-like structures (arrow). (E) Frequency and expression (MFI) of CD27, CD244 and NKp30 on NK cells cultured alone (NK) or after 18 hours of coculture with CD14+CD16− monocytes (NK + Mo). (F) Distribution of IFN-γ during the interaction of NKp30-blocked NK cells with monocytes. (G) IFN-γ dynamics during the interaction of NKp30-blocked NK cells with monocytes. (H) Total fluorescence intensity of IFN-γ determined at the junction formed between NK cells and monocytes. Prestained NK cells (green) and monocytes (violet) were cocultured and live-cell imaging was recorded for up to 12 hours. (I) Captures of the interaction between NK cells and monocytes. (J) Interaction time of monocytes with NK cells vs monocytes with NKp30-blocked NK cells. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001. See “Confocal microscopy” for stains used; original magnifications ×20 (A-B,F,I) and ×40 (C-D,G).
Among NK-cell receptors that are known to recognize cognate ligands on monocytes, NKp30 (but neither CD27 or CD224) was upregulated in NK cells after cell-to-cell contact with monocytes (Figure 3E) and its expression was sustained by soluble factors produced by monocytes (supplemental Figure 2B). Across groups, no significant differences on the B7-H6 expression on monocytes were found (supplemental Figure 2C). The blockage of NKp30 during NK cells and monocytes interaction abrogates the polarization of IFN-γ toward the junction (Figure 3F,H) and the IFN-γ distribution within the cells resembles that observed in nonagglomerated NK cells (Figure 3B,G). Live-cell imaging revealed that when NKp30 is blocked, interactions between cells are less frequent and shorter (Figure 3I-J).
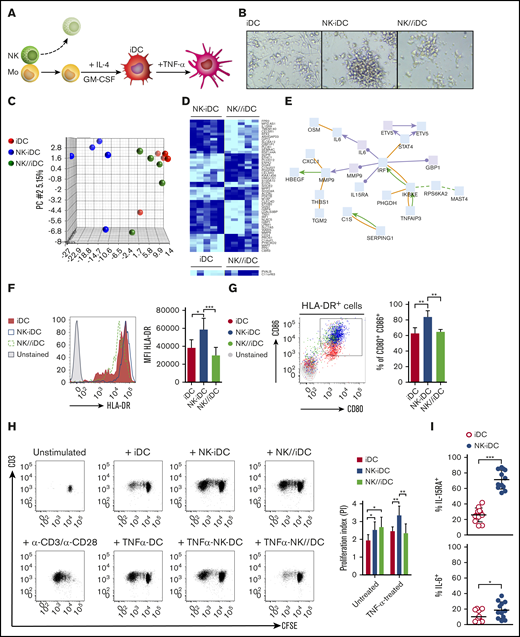
DCs derived from monocytes instructed by direct contact with NK cells retain a stable transcriptional program and present enhanced antigen-presenting capacity
To investigate whether NK cells induced stable changes in monocytes, we removed NK cells from the culture and induced differentiation of monocytes into DCs (Figure 4A). Immature DCs derived from monocytes previously cocultured in direct contact with NK cells (NK-iDCs) formed large clusters unlike those derived from control monocytes (iDCs) or from monocytes exposed to NK cells through a Transwell membrane (NK//iDCs) (Figure 4B). Principal component analysis (PCA) of the gene-expression profile revealed that NK-iDCs have a distinct transcriptional program from that of NK//iDCs and iDCs (Figure 4C). In NK-iDCs, 51 genes were differentially expressed (P < .005) compared with NK//iDCs (Figure 4D). NK//iDCs and iDCs shared a similar gene-expression signature, closely clustering in the PCA and exhibiting differences in only 2 genes. Changes in monocytes and Mo-DCs persisted only when induced by cell-to-cell contact with NK cells (supplemental Table 2). Network analysis29 of differentially expressed genes revealed a persistent IRF1 signature as the main transcription factor activated in NK-Mo and its subsequent derived NK-iDCs (Figure 4E; supplemental Figure 3), orchestrating in DCs the upregulation of genes important for migration (MMP9), antiviral activity (GBP1, IKBKE), IL-12 production and Th1 polarization (STAT4), Th17 polarization (IL6, CXCL1), and NK and CD8+ T cells survival and proliferation (IL15RA) (Figure 4E; supplemental Figure 4A-B).

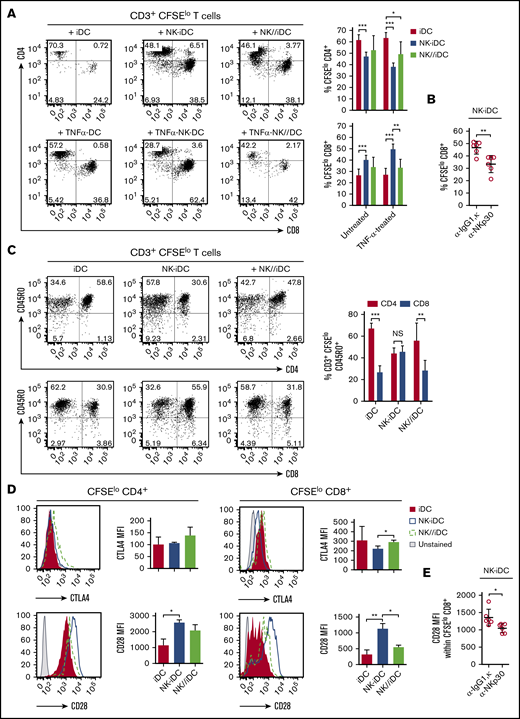
DCs derived from monocytes primed by cell-to-cell contact with NK cells display a unique transcriptional program and enhanced antigen-presenting capacity. (A) Immature DCs derived from NK-cell–primed CD14+CD16− monocytes were obtained after NK-cell depletion and 5-day treatment with IL-4 and GM-CSF (NK-iDCs). Immature DCs derived from monocytes alone (iDCs) or primed by NK cells through a Transwell membrane (NK//iDCs) were used as controls. For some experiments, immature DCs were activated for 24 hours using TNF-α. (B) DCs at fifth day of differentiation, photographed at an original magnification of ×20 using an inverted microscope. After differentiation, iDCs were sorted by flow cytometry and total RNA was extracted for the determination of gene expression by microarray. (C) Transcriptional heterogeneity between DCs determined by principal component analysis (PCA) of differentially expressed genes (P < .005). (D) Heatmaps for differentially expressed genes (P < .005) of DCs derived from monocytes primed by NK cells through cell-cell contact (NK-iDCs) vs Transwell contact (NK//iDCs); control iDCs vs Transwell contact (NK//iDCs). (E) Transcriptional network upregulated in DCs derived from NK cell-primed monocytes (NK-iDCs). After differentiation, the surface phenotype of iDCs was analyzed by flow cytometry. (F) HLA-DR histograms and expression (MFI) of iDCs at different conditions. (G) Dot plot and frequency of CD86+ CD80+ iDCs at different conditions. Untreated and TNF-α–treated iDCs from different conditions were cocultured with purified, CFSE-stained naive allogeneic T cells for 5 days. Unstimulated T cells and T cells stimulated with α-CD3/α-CD28 beads were used as negative and positive proliferation controls, respectively. (H) Dot plots showing total CD3 T proliferation (CFSE dilution) induced by DCs from different conditions by the fifth day, assessed by flow cytometry; proliferation index obtained in each condition determined in FlowJo. (I) Frequency of IL-15RA+ and IL-6+ DCs derived from NK-primed monocytes, assessed by flow cytometry after 48 hours of coculture with purified naive allogeneic T cells. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001.
DCs derived from monocytes primed by cell-to-cell contact with NK cells display a unique transcriptional program and enhanced antigen-presenting capacity. (A) Immature DCs derived from NK-cell–primed CD14+CD16− monocytes were obtained after NK-cell depletion and 5-day treatment with IL-4 and GM-CSF (NK-iDCs). Immature DCs derived from monocytes alone (iDCs) or primed by NK cells through a Transwell membrane (NK//iDCs) were used as controls. For some experiments, immature DCs were activated for 24 hours using TNF-α. (B) DCs at fifth day of differentiation, photographed at an original magnification of ×20 using an inverted microscope. After differentiation, iDCs were sorted by flow cytometry and total RNA was extracted for the determination of gene expression by microarray. (C) Transcriptional heterogeneity between DCs determined by principal component analysis (PCA) of differentially expressed genes (P < .005). (D) Heatmaps for differentially expressed genes (P < .005) of DCs derived from monocytes primed by NK cells through cell-cell contact (NK-iDCs) vs Transwell contact (NK//iDCs); control iDCs vs Transwell contact (NK//iDCs). (E) Transcriptional network upregulated in DCs derived from NK cell-primed monocytes (NK-iDCs). After differentiation, the surface phenotype of iDCs was analyzed by flow cytometry. (F) HLA-DR histograms and expression (MFI) of iDCs at different conditions. (G) Dot plot and frequency of CD86+ CD80+ iDCs at different conditions. Untreated and TNF-α–treated iDCs from different conditions were cocultured with purified, CFSE-stained naive allogeneic T cells for 5 days. Unstimulated T cells and T cells stimulated with α-CD3/α-CD28 beads were used as negative and positive proliferation controls, respectively. (H) Dot plots showing total CD3 T proliferation (CFSE dilution) induced by DCs from different conditions by the fifth day, assessed by flow cytometry; proliferation index obtained in each condition determined in FlowJo. (I) Frequency of IL-15RA+ and IL-6+ DCs derived from NK-primed monocytes, assessed by flow cytometry after 48 hours of coculture with purified naive allogeneic T cells. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001.
Unlike iDCs or NK//iDCs, NK-iDCs show a single peak of HLA-DRhigh expression (Figure 4F) and preferentially expressed the IFN-γ receptor 1 (CD119) (supplemental Figure 4C). CD80 and CD86 were coexpressed at higher density and in a larger proportion of cells compared with iDCs (Figure 4G; supplemental Figure 4D). NK-iDCs induced higher proliferation of allogeneic T cells and its activation with TNF further increased that ability (Figure 4H). During DC and T-cell cocultures and consistently with gene-expression data, a higher frequency of IL-15RA+ and IL-6+ DCs was found within NK-iDCs compared with iDCs (Figure 4I; supplemental Figure 4D-E). A modest upregulation of IL-23+ and IL-26+ DCs were also found in NK-iDCs compared with iDCs (supplemental Figure 4E).
DCs derived from monocytes instructed by direct contact with NK cells preferentially stimulate CD8+ T-cell proliferation
NK-iDCs are enriched with cells exhibiting distinctive transcriptional and functional profiles. After coculture with allogeneic naive CD45RA+CD3+ T cells, NK-iDCs preferentially induced the activation and proliferation of CD8+ T cells (Figure 5A) and CD45RO+CD8+ T cells (Figure 5C), whereas iDCs and NK//iDCs preferentially induced the proliferation of CD4+ T cells and CD45RO+CD4+ T cells; the activation of DCs with TNF further intensifies these phenomena (Figure 5A). The CD8+ T cells stimulated by NK-iDCs presented low expression of the inhibitory molecule CTLA-4 and high expression of the activating molecule CD28 (Figure 5D). The CD8+ T-cell proliferation and activation bias of NK-iDCs is abrogated by the blockage of NKp30, during NK-cell and monocyte interaction (Figure 5B,E).
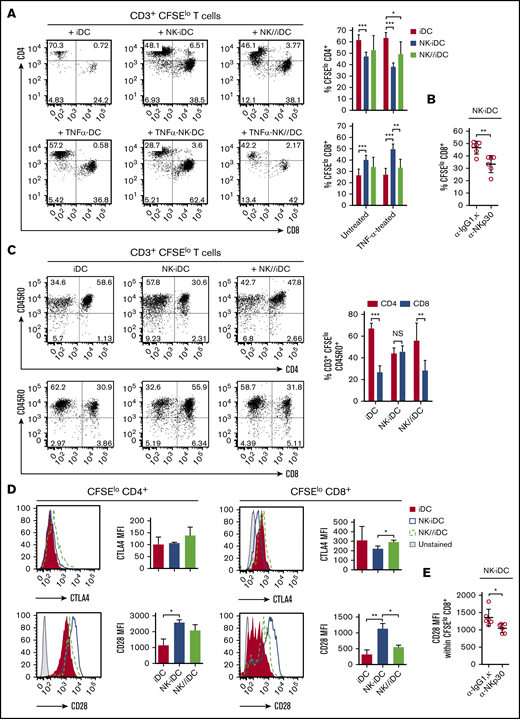
DCs derived from NK-cell–primed monocytes through cell-to-cell contact via NKp30 promote CD8+T-cell activation and proliferation. Untreated and TNF-α–treated iDCs derived from NK-cell primed monocytes (NK-iDCs) were cocultured with purified, CFSE-stained naive allogeneic T cells for 5 days. DCs derived from monocytes alone (iDCs) or primed by NK cells through a Transwell membrane (NK//iDCs) were used as controls. By the fifth day of coculture, the phenotypical and functional profiles of T cells were assessed by flow cytometry. (A) Frequency of proliferating (CFSElow) CD4+ T cells and CD8+ T cells stimulated by DCs at different conditions. (B) Frequency of proliferating (CFSElow) CD8+ T cells stimulated by DCs derived from monocytes primed by NK cells in the presence of anti-NKp30 (α-NKp30) or IgG1,κ isotype control (IgG1,κ). (C) Frequency of proliferating (CD3+CFSElow) CD45RO+CD4+ T cells and CD8+ T cells stimulated by DCs at different conditions. (D) Expression (MFI) of CTLA-4 and CD28 by proliferating (CFSElow) CD4+ T cells and CD8+ T cells stimulated by DCs at different conditions. (E) Expression (MFI) of CD28 by proliferating (CFSElo) CD8+ T cells stimulated by DCs derived from monocytes primed by NK cells in the presence of anti-NKp30 (α-NKp30) or IgG1,κ isotype control (IgG1,κ). Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001.
DCs derived from NK-cell–primed monocytes through cell-to-cell contact via NKp30 promote CD8+T-cell activation and proliferation. Untreated and TNF-α–treated iDCs derived from NK-cell primed monocytes (NK-iDCs) were cocultured with purified, CFSE-stained naive allogeneic T cells for 5 days. DCs derived from monocytes alone (iDCs) or primed by NK cells through a Transwell membrane (NK//iDCs) were used as controls. By the fifth day of coculture, the phenotypical and functional profiles of T cells were assessed by flow cytometry. (A) Frequency of proliferating (CFSElow) CD4+ T cells and CD8+ T cells stimulated by DCs at different conditions. (B) Frequency of proliferating (CFSElow) CD8+ T cells stimulated by DCs derived from monocytes primed by NK cells in the presence of anti-NKp30 (α-NKp30) or IgG1,κ isotype control (IgG1,κ). (C) Frequency of proliferating (CD3+CFSElow) CD45RO+CD4+ T cells and CD8+ T cells stimulated by DCs at different conditions. (D) Expression (MFI) of CTLA-4 and CD28 by proliferating (CFSElow) CD4+ T cells and CD8+ T cells stimulated by DCs at different conditions. (E) Expression (MFI) of CD28 by proliferating (CFSElo) CD8+ T cells stimulated by DCs derived from monocytes primed by NK cells in the presence of anti-NKp30 (α-NKp30) or IgG1,κ isotype control (IgG1,κ). Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction. *P < .05, **P < .01, ***P < .001.
DCs derived from monocytes instructed by direct contact with NK cells drive the polarization of Tc17/IFN-γ cells
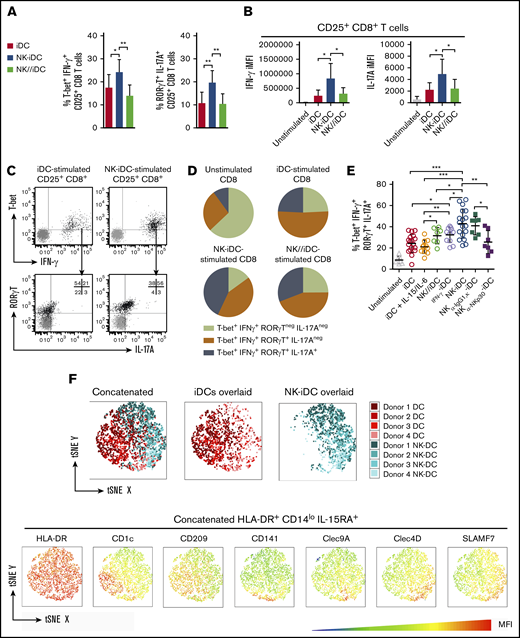
Activated CD8+ T cells induced by NK-iDCs displayed higher frequency of T-bet+IFN-γ+ (T cytotoxic type 1 [Tc1]) and RORγT+IL-17A+ (T cytotoxic type 17 [Tc17]) cells (Figure 6A) and higher cytoplasmic level of IFN-γ and IL-17A, respectively, than those induced by iDCs and NK//iDCs (Figure 6B). These data are consistent with the transcriptional program displayed by NK-iDCs (Figure 4E) and their cytokine production profile (Figure 4I). Because some Tc1 cells express the transcription factor RORγT (Figure 6C), we investigated whether NK-DCs induce a CD8+ T-cell population able to simultaneously produce IFN-γ and IL-17A. iDCs and NK//iDCs preferentially stimulated a T-bet+RORγT+CD8+ T-cell population, able to produce IFN-γ but less frequently IL-17A. In contrast, NK-iDCs significantly enhanced the production of both IFN-γ and IL-17A from T-bet+RORγT+CD8+ T cells (Figure 6D). Treating monocytes with IFN-γ before differentiation increased in DCs the capacity to polarize toward Tc17/IFN-γ, however, NK-DCs were superior; blocking the NKp30 receptor during the interaction of NK cells and monocytes weakened the capacity of NK-iDCs to polarize naive CD8+ T cells toward Tc17/IFN-γ cells (Figure 6E). Both NK cells subtypes (CD56bright and CD56low) were required to prime DC precursors that stimulate IFN-γ production by CD8+ T cells whereas CD56low alone, but not CD56bright, enhanced the ability of DC to sustain CD8+ T-cell proliferation (supplemental Figure 5A-B).
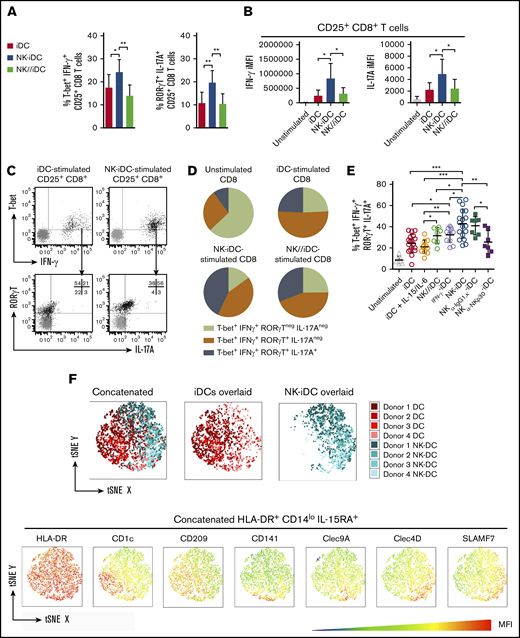
DCs derived from NK-cell–primed monocytes through cell-to-cell contact via NKp30 drive the polarization of IFN-γ and IL-17A-producers type 17 CD8+T cells. DCs derived from NK-cell–primed monocytes (NK-iDCs) were cocultured with purified naive allogeneic T cells for 5 days. DCs derived from monocytes alone (iDCs) or primed by NK cells through a Transwell membrane (NK//iDCs) were used as controls. By the fifth day of coculture, T-cell phenotype and capacity to produce cytokines after PMA plus ionomycin stimuli was assessed by flow cytometry. (A) Frequency of T-bet+IFN-γ+ and RORγT+IL-17A+ within CD25+CD8+ T cells stimulated by DCs at different conditions. (B) Production (iMFI: frequency multiplied by MFI) of IFN-γ and IL-17A by CD25+CD8+ T cells stimulated by DCs at different conditions. (C) Gating strategy for the analysis of polarized CD25+CD8+ T cells. (D) Comparison of different types of CD25+CD8+ T cells polarized by DCs from different conditions. (E) Frequency of IFN-γ and IL-17A producers type 17 CD8+ T cells (T-bet+IFN-γ+RORγT+IL-17A+) stimulated by control DCs and DCs derived from monocytes primed by NK cells, in the presence of anti-NKp30 (α-NKp30) or IgG1,κ isotype control (IgG1,κ). (F) tSNE analysis of concatenated samples (all conditions combined) and split by iDCs and NK-iDCs overlaid samples, respectively; MFI statistic heatmaps of the concatenated sample for DC markers. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welchs correction. *P < .05, **P < .01, ***P < .001.
DCs derived from NK-cell–primed monocytes through cell-to-cell contact via NKp30 drive the polarization of IFN-γ and IL-17A-producers type 17 CD8+T cells. DCs derived from NK-cell–primed monocytes (NK-iDCs) were cocultured with purified naive allogeneic T cells for 5 days. DCs derived from monocytes alone (iDCs) or primed by NK cells through a Transwell membrane (NK//iDCs) were used as controls. By the fifth day of coculture, T-cell phenotype and capacity to produce cytokines after PMA plus ionomycin stimuli was assessed by flow cytometry. (A) Frequency of T-bet+IFN-γ+ and RORγT+IL-17A+ within CD25+CD8+ T cells stimulated by DCs at different conditions. (B) Production (iMFI: frequency multiplied by MFI) of IFN-γ and IL-17A by CD25+CD8+ T cells stimulated by DCs at different conditions. (C) Gating strategy for the analysis of polarized CD25+CD8+ T cells. (D) Comparison of different types of CD25+CD8+ T cells polarized by DCs from different conditions. (E) Frequency of IFN-γ and IL-17A producers type 17 CD8+ T cells (T-bet+IFN-γ+RORγT+IL-17A+) stimulated by control DCs and DCs derived from monocytes primed by NK cells, in the presence of anti-NKp30 (α-NKp30) or IgG1,κ isotype control (IgG1,κ). (F) tSNE analysis of concatenated samples (all conditions combined) and split by iDCs and NK-iDCs overlaid samples, respectively; MFI statistic heatmaps of the concatenated sample for DC markers. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welchs correction. *P < .05, **P < .01, ***P < .001.
Unlike CD8+ T cells, NK-iDCs did not polarize naive CD4+ T cells toward a Th1, Th2, or Th17 phenotype (supplemental Figure 5C). However, NK-iDCs presented an enhanced capacity to trigger IL-21 production by the stimulated CD25+CD4+ T cells, prevented by the blockage of NKp30 during NK-cell and monocyte interaction (supplemental Figure 5D).
t-distributed Stochastic Neighbor Embedding (tSNE) analysis of flow cytometry data revealed major phenotypic changes within HLA-DR+CD14lowIL-15RA+ DCs. NK-iDCs were enriched in cells expressing the endocytic receptor Clec4D (CLECSF8/Dectin-3) and the costimulatory molecule SLAMF7 (CD319) (Figure 6F). CD1c+ and CD209+ cells were enriched within iDCs, whereas CD141+ and Clec9A+ cells were similarly distributed within NK-iDCs and iDCs. Preliminary data from an acute myeloid leukemia (AML) patient treated with an NK-cell–based immunotherapy (NK-IT) showed that the frequency of Clec4D+SLAMF7+ cells within HLA-DR+CD14lowIL-15RA+ PBMCs were upregulated after NK-IT treatment and in comparison with age-matched HDs (supplemental Figure 6).
Tc17/IFN-γ response is decreased in patients with classical NK-cell deficiency
To understand the in vivo relevance of the phenomena hereby described, we investigated the populations of DCs and CD8+ T cells circulating in the blood of patients with classical NK-cell deficiency driven by mutations in the GATA2 gene. We examined the frequency of CD3−CD56+ NK cells and Lin−CD14+ monocytes in the PBMCs of 7 patients (Figure 7A). For 1 of them, we performed single-cell sequencing analysis before and after bone marrow transplant (Figure 7B), which revealed that before the transplant, myeloid cells lack the transcriptional signature that characterized NK-primed monocytes and Mo-DCs (Figure 7C). The subset of HLA-DR+CD14lowIL-15RA+ DCs, identified in vitro as NK-cell primed, was significantly downregulated in PBMCs of GATA2-deficient patients; within that DC subset, Clec4D+SLAMF7+ DCs, as well as CD141+Clec9A+ DCs, were significantly decreased in patients with GATA2 deficiency (Figure 7D). Monocle analyses34 of myeloid cells show that either before and after transplant, they were able to undergo further differentiation (supplemental Figure 7A).
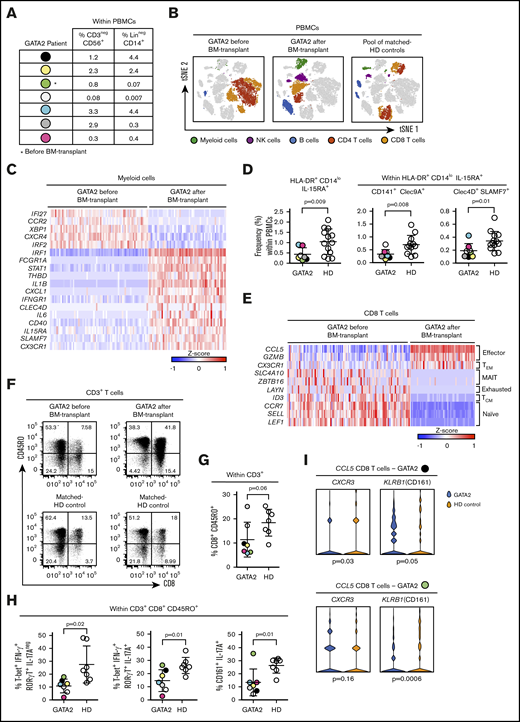
Patients with classical NK-cell deficiency (GATA2 mutations) have low frequencies of Clec4D+SLAMF7+DCs and reduced Tc17/ IFN-γ responses. PBMCs derived from blood of 7 patients bearing mutations on the GATA2 gene were analyzed by flow cytometry. PBMCs from 2 patients (indicated by black and green dots) were analyzed at a single-cell level; only for 1 patient (green dot), samples before and after bone marrow transplant were obtained; PBMCs from HD were used as controls. (A) Frequencies of CD3−CD56+ (NK cells) and Lin−CD14+ cells (monocytes) in PBMCs from patients with GATA2 deficiency. (B) tSNE analysis of the different populations found in PBMCs before and after transplant. (C) Heatmap of genes differentially expressed on the population of myeloid cells, before and after transplant. (D) Frequency of HLA-DR+CD14lowIL-15RA+ DCs from GATA2 patients and HD controls. (E) Heatmap of genes differentially expressed on the population of CD8 T cells, before and after transplant. (F) Dot plots of CD45RO+CD8+ T cells from a GATA2 patient before and after transplant with the corresponding HD control. (G) Frequency of CD45RO+CD8+ T cells from GATA2 patients and HD controls. PBMCs from GATA2 patients and HDs were stimulated for 6 hours by PMA and ionomycin. (H) Frequency of Tc1, Tc17/IFN-γ and Tc17 cells from GATA2 patients and HD controls. (I) Gene expression of CXCR3 and KLRB1 on CCL5 expressing CD8 T cells from GATA2 patients and HD controls. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction.
Patients with classical NK-cell deficiency (GATA2 mutations) have low frequencies of Clec4D+SLAMF7+DCs and reduced Tc17/ IFN-γ responses. PBMCs derived from blood of 7 patients bearing mutations on the GATA2 gene were analyzed by flow cytometry. PBMCs from 2 patients (indicated by black and green dots) were analyzed at a single-cell level; only for 1 patient (green dot), samples before and after bone marrow transplant were obtained; PBMCs from HD were used as controls. (A) Frequencies of CD3−CD56+ (NK cells) and Lin−CD14+ cells (monocytes) in PBMCs from patients with GATA2 deficiency. (B) tSNE analysis of the different populations found in PBMCs before and after transplant. (C) Heatmap of genes differentially expressed on the population of myeloid cells, before and after transplant. (D) Frequency of HLA-DR+CD14lowIL-15RA+ DCs from GATA2 patients and HD controls. (E) Heatmap of genes differentially expressed on the population of CD8 T cells, before and after transplant. (F) Dot plots of CD45RO+CD8+ T cells from a GATA2 patient before and after transplant with the corresponding HD control. (G) Frequency of CD45RO+CD8+ T cells from GATA2 patients and HD controls. PBMCs from GATA2 patients and HDs were stimulated for 6 hours by PMA and ionomycin. (H) Frequency of Tc1, Tc17/IFN-γ and Tc17 cells from GATA2 patients and HD controls. (I) Gene expression of CXCR3 and KLRB1 on CCL5 expressing CD8 T cells from GATA2 patients and HD controls. Error bars represent standard deviation. Data are representative of at least 2 or more independent experiments combined; n = 2-3 HDs per experiment. Statistical comparisons were performed with the unpaired Student t test with Welch correction.
Before the bone marrow transplant, the CD8+ T-cell population mainly consisted of naive cells (Figure 7E) whereas CD8+CD45RO+ T cells were less frequent than in HDs or after transplant (Figure 7F). Most GATA2-deficient patients had a lower frequency of CD8+CD45RO+ T cells than all the tested HDs (Figure 7G). After phorbol 12-myristate 13-acetate (PMA) plus ionomycin stimulation, decreased Tc1 and Tc17 responses were observed in GATA2-deficient patients, compared with HDs (Figure 7H). However, innate cell alterations variability among patients (Figure 7A) might be associated with additional defects or compensatory mechanisms regulating the circulating CD8+ T subset, suggesting caution in data interpretation. For instance, a GATA2-deficient patient (black dot) with low blood frequency of NK cells and close-to-normal frequency of monocytes has low CXCR3 (Tc1) and KLRB1 (Tc17) CCL5+CD8+ T cells; whereas a GATA2-deficient patient (green dot) with low frequencies of both NK cells and monocytes, has low KLRB1 CCL5+CD8+ T cells and mucosal-associated invariant T (MAIT) cells within the CD8 T cells subset, possibly expanded as a compensatory mechanism (supplemental Figure 7B).
Discussion
Human Tc17 cells comprise a small population of circulating CD8+ T cells that originate from naive CD45RA+ precursors and are predominantly found within the CD28+CD45RA− memory compartment.35 Tc17 cells retain a functional plasticity that allows them to produce IFN-γ in addition to IL-1736 ; this ability has been positively correlated with the exacerbation of pulmonary inflammation, the protection against lethal influenza, and the regression of large established B16F10 melanomas.37-39 Tc17 cells have been studied in the context of skewed in vitro differentiation using IL-6 and TGF-β; what drives their polarization during an immune response remains unknown. Our data suggest that NK cells drive Tc17/IFN-γ cells through priming of Mo-DCs. This priming takes place during early encounters between activated NK cells and monocyte precursors of DCs and imprint a functional specialization that, via IL-15 and IL-6 production, supports the differentiation of Tc17 cells and, probably, of IL-21–producing CD4+ T cells (Th21). IL-21 has been shown to promote durable CD8+ T-cell activation, survival, as well as to favor the generation of memory CD8+ T-cell responses.40,41 Despite being widely recognized that naive T-cell polarization rely on signals provided during antigen presentation, our results suggest that Tc17-mediated immunity might evolve and be tuned by NK cells during the early events of an immune response.
The critical element connecting NK cells to Tc17 cells are Mo-DCs. During cocultures of NK cells and monocyte, differential exposure to IFN-γ at early stages of the differentiation process (in terms of spatial, temporal, and gradient exposure) likely resulted in different signaling outcomes, introducing variability to what is thought to be a “homogeneous” DC population. The identification of NK-cell–instructed Mo-DCs was challenging, once this population is diluted within a mixed population of DCs with partially overlapping markers. Still, NK-iDCs were characterized by a predominance of cells expressing Clec4D and CD319 whereas DCs expressing CD1c and CD209 are scarce. Clec4D, a C-type lectin receptor important for phagocytosis and proinflammatory-cytokine production by myeloid cells, is essential for immunity against Mycobacterium tuberculosis and Candida albicans.42,43 In turn, CD319 is upregulated in plasmacytoid DCs (pDCs) of systemic lupus erythematosus (SLE) patients, characterized by increased capacity to produce type I IFNs when stimulated with immune complexes in the presence of NK cells.44 Microarray analysis of both monocytes and Mo-DCs highlights a novel phenomenon resembling trained immunity, defined as the increased inflammatory responsiveness of innate cells, resulting in an amplified response and resistance to subsequent nonspecific challenges.45,46 The stable transcriptomic changes in monocytes that persisted during the differentiation in DC strongly suggest a stable epigenetic mechanism induced by NK cells.
It is accepted that cell-to-cell contact between NK cells and myeloid cells is dispensed during their cytokine cross talk. However, intercellular communication through cytokines depends on the characteristics of cytokine niches within cellular microenvironments, in which the strength of the signaling induced in the cytokine-consuming cells depends on its distance from the producer.47-49 Here, we show that the distance between NK cells and monocytes drastically impacted IFN-γ signaling. Moreover, the participation of the NKp30 receptor is crucial for the acquisition of the transcriptional program and functional specialization of Mo-DCs and directly affects the localization of IFN-γ at the synapse between monocytes and NK cells. Besides its role as an activator receptor that leads to the lysis of B7-H6+ cells, NKp30 is crucial for the stabilization of NK-cell synapses.50,51 Here, we show that the maintenance of long synapses via NKp30 promoted the polarization of IFN-γ, required for imprinting monocytes with their stable, functional specialization.
The killing of a subset of monocytes is a complex process that involves not only the engagement of NKp30 but also an imbalance of many signals leading to lysis, such as low expression of MHC class I molecules and high expression of B7-H6. Yet, the elimination of a monocyte population by itself is not sufficient for the functional specialization of Mo-DCs; rather, NK cells directly license and instruct the remaining monocytes. The balance between NK cells ability to train or kill monocytes modulates immunity. It has been shown that the lack of perforin in NK-cell synapses with myeloid cells prevents cell-to-cell disengagement, leading to repetitive calcium signaling that amplifies the amount of inflammatory cytokines secreted by both NK and myeloid cells.52 Decreased numbers of NK cells with reduced cytotoxic function and increased capacity to produce IFN-γ are found in blood from patients with active autoimmune diseases.53-56 Yet, NK cells comprise one of the most prominent populations found in inflamed joints of patients with rheumatoid arthritis (RA), colocalizing with monocytes, DCs, and T cells.10,57,58 Through the production of GM-CSF, the CD56bright NK-cell subset derived from patients’ synovial fluid is capable to trigger and maintain the in vitro differentiation of CD14+ monocytes into “DC-like” cells.10 CD14+ monocytes and inflammatory DCs found in synovial fluids from patients with RA potently induce the in vitro polarization and expansion of Th17 cells,59-61 which are largely involved in the development of chronic inflammation and severity of RA and other autoimmune and inflammatory diseases.62,63 It is plausible that, by promoting the priming and survival of inflammatory monocytes and DCs that sustain Th17 and Tc17 activation, NK cells may play an early and important role in the pathogenesis and maintenance of autoimmunity.
It is likely that both the killing and instruction of monocytes by NK cells is impaired in patients with the classical NK-cell deficiency (cNKD). At baseline, cNKD patients share no other immune aberrations such as defects in the cytotoxic machinery shared with CTLs.64 The fact that protective CTL responses against intracellular pathogens and tumors cannot be mounted properly in these patients is intriguing. NK cells play an important, although not exclusive, role in the supply of antigenic debris and cytokine milieu that enhance DCs’ microbial and tumoral antigen presentation and cross-presentation to Th cells and CTLs.6,7,15,65-67 Still, additional mechanisms are linking NK cells to CD8+ T-cell–mediated responses. Although NK-cell numbers may vary in patients with GATA2 mutations from almost absent to nearly normal, their capacity for lytic function and cytokine production is impaired.68-70 Although B-cell, DC, and NK-cell cytopenias are most commonly described in GATA2 deficiency, T-cell abnormalities have also been described but the mechanism by which they occur is unknown.71-74 Here, we show that loss of NK-cell numbers and function parallels a decreased presence of NK-cell–primed Mo-DCs, with a concomitant impairment in Tc1, Tc17/IFN-γ, and Tc17 responses. Notably, following hematopoietic stem cell transplant, the gene signature of NK-Mo/NK-DCs and memory CD8 T cells found in vitro are restored in the blood of one patient; moreover, the Clec4D+SLAMF7+ NK-DC population found in vitro is upregulated in blood of an AML patient after the infusion of NK cells, suggesting that reconstitution of a functional NK-cell compartment might restore monocytes, DCs, and CD8 T-cell functionality, perhaps with the participation of the mechanism herein described. To our knowledge, our findings represent the first evidence connecting NK cells to the development of Tc17 cells through Mo-DCs. This new information has implications in the understanding of Tc17-mediated immune responses in humans and could be exploited for immunotherapy contexts in which IFN-γ and IL-17A production by CD8+ T cells may be either boosted or targeted.
The microarray data reported in this article have been deposited in the Gene Expression Omnibus database (accession number GSE116487).
For original data, please contact mariasalomon@usp.br.
Acknowledgments
The authors thank Robin Winkler-Pickett and Sharon Bargo for administrative and logistical support in the development of this project. The authors thank Karen M. Wolcott, Langston Lim, Jonathan H. Badger, Michael Kelly, and Carolina C. Salomón for assistance in cell sorting, live-cell imaging, bioinformatic analyses, single-cell sequencing, and scientific illustration, respectively. The authors appreciate the support from the staff of the NIH Clinical Center Department of Transfusion Medicine. The authors thank Diane Cooper, NIH Library Writing Center, for manuscript editing assistance.
This work was supported by the Intramural Research Program of the National Institutes of Health (National Cancer Institute, Center for Cancer Research, Cancer and Inflammation Program, grant 1ZIABC011152) (G.T.); Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; grant 2009/54599-5 [J.A.M.B.] and grants 2011/05331-0, 2012/23478-0, 2014/10290-9, and 2017/13686-9 [M.A.C.-S.]); Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (J.A.M.B.); National Institutes of Health, National Institute of Allergy and Infectious Diseases grant R01AI120989 (J.S.O.); and National Institutes of Health, National Institute of Allergy and Infectious Diseases grant R01AI137275 and an American Society for Hematology Junior Scholar award (E.M.M.).
The authors acknowledge the contribution and collaboration of Steven Holland and Eugene Frenkel (deceased) who provided clinical care and obtained patient samples from the GATA2-deficient patients.
Authorship
Contribution: M.A.C.-S. designed the research, conducted experiments, acquired and analyzed the data together with R.S. (gene expression), S.R. (confocal microscopy), R.X.d.N. (gene expression and single cell), A.D. (single cell), H.S.-C. (cell isolation and culture, gene expression and ELISA), K.S.-C.B. (live cell imaging), J.S.O. and E.M.M. (GATA2 deficiency), and L.S. (NK-cell–based immunotherapy); M.A.C.-S. wrote and revised the manuscript; J.A.M.B. and G.T. supervised the project and reviewed the manuscript along with R.S., H.S.-C., R.X.d.N., K.S.-C.B., E.M.M., and A.D.; and all authors approved the final draft of the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
The current affiliation for H.S.-C. is Institute of Tropical Pathology and Public Health, Federal University of Goias, Goiania, Brazil.
Correspondence: Maria A. Clavijo-Salomon, Center of Translational Research in Oncology, Institute of Cancer of São Paulo (ICESP), University of São Paulo Medical School, São Paulo, Brazil; e-mail: mariasalomon@usp.br; José A. M. Barbuto, Cell and Molecular Therapy Center (NETCEM), University of São Paulo, São Paulo, Brazil; e-mail: jbarbuto@icb.usp.br; and Giorgio Trinchieri, National Cancer Institute, Building 37, Room 4146A, Bethesda, MD 21702-1201; e-mail: trinchig@niaid.nih.gov.