

In this issue of Blood, reveal a previously unknown role for the canonical Wnt signaling pathway in the regulation of granulocyte production in both steady-state and during emergency granulopoiesis.1
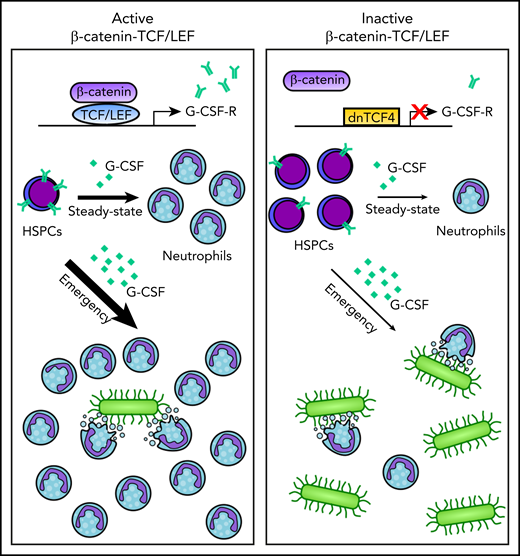
Active β-catenin-TCF/LEF signaling drives expression of the G-CSF receptor on HSPCs which, in turn, renders HSPCs more responsive to G-CSF thereby promoting steady-state and emergency granulopoiesis (left panel). By contrast, disruption of the β-catenin-TCF/LEF signaling results in impaired steady-state and emergency granulopoiesis (right panel). See graphical abstract in the article by Danek et al that begins on page 2574.
Active β-catenin-TCF/LEF signaling drives expression of the G-CSF receptor on HSPCs which, in turn, renders HSPCs more responsive to G-CSF thereby promoting steady-state and emergency granulopoiesis (left panel). By contrast, disruption of the β-catenin-TCF/LEF signaling results in impaired steady-state and emergency granulopoiesis (right panel). See graphical abstract in the article by Danek et al that begins on page 2574.
Neutrophilic granulocytes serve as a first line of defense against pathogens. Adequate numbers are critical for survival, as demonstrated by the severe and often fatal infection in patients with congenital or iatrogenic neutropenia.2 Given their short half-life, neutrophils need to be constantly replenished to maintain steady-state neutrophil counts. Moreover, upon severe systemic infection, when neutrophils are being consumed, granulopoietic output is massively enhanced to meet the high demand. Consequently, an intricate and redundant regulatory network consisting of cytokines/growth factors, their cognate receptors, and downstream transcriptional programs has evolved to sustain appropriate, demand-adapted granulocyte numbers.3
Danek et al now identify the canonical Wnt signaling pathway as yet another important regulator of granulopoiesis. The canonical Wnt pathway signals via various secreted Wnt ligands that bind to the Frizzled receptors, leading to stabilization and accumulation of β-catenin and its translocation to the nucleus, where it ultimately acts as transcriptional coactivator by interacting with members of the T-cell factor/lymphoid enhancer-binding factor (TCF/LEF) family of transcription factors such as TCF4.4 The role of the canonical Wnt signaling pathway in hematopoietic stem and progenitor cell (HSPC) biology has been extensively studied, yielding conflicting results.5 Stimulatory, inhibitory or even neutral effects have been described, and these discrepancies are likely due to use of different experimental model systems that interfere with canonical Wnt signaling at various levels of the pathway and to varying degrees.6
In an attempt to overcome these likely limitations, Danek et al use a novel mouse model that employs a dominant-negative form of TCF4 (dnTCF4) whereby transcriptional activity of β-catenin-TCF4 in HSPCs is impaired, whereas the nontranscriptional activities of β-catenin as well as noncanonical Wnt signaling are left intact. The authors show that mice expressing dnTCF4 have reduced peripheral blood and bone marrow (BM) neutrophils and increased immunophenotypically defined HSPC subsets with the exception of long-term hematopoietic stem cells HSCs (LT-HSCs). This accumulation of HSPCs is due to impaired granulocytic differentiation. To elucidate the mechanistic basis for the observed differentiation defect of dnTCF4-expressing HSPCs, gene expression analyses were performed and compared with WT HSPCs, thereby identifying a number of genes known to be important in regulating granulopoiesis. Among these downregulated genes in dnTCF4-expressing HSPCs is Csf3r, encoding the receptor for granulocyte colony-stimulating factor (G-CSF), a master regulator of steady-state and emergency granulopoiesis. Accordingly, cell surface G-CSF receptor (G-CSF-R) expression is reduced on murine HSPCs expressing dnTCF4. These findings suggest that the lack of β-catenin-TCF4–mediated transcription of Csf3r and the resulting reduced expression of the G-CSF-R is responsible for the observed defect in granulocytic differentiation (see figure). Indeed, Danek et al provide evidence that TCF/LEF transcription factors directly bind to the CSF3R promoter and enhancer in a number of human cell lines, and importantly, that treatment with G-CSF results in reduced downstream JAK/STAT3 signaling and granulocytic differentiation in dnTCF4-expressing HSPCs.
Intriguingly, the fact that mice deficient in the master transcriptional regulator of steady-state hematopoiesis, CCAAT enhancer binding protein α (C/EBPα), completely lack granulocyte differentiation,7 demonstrates that β-catenin-TCF4–mediated transcription fails to compensate for C/EBPα deficiency, and thus primarily acts by fine-tuning steady-state granulopoiesis. However, the striking similarities between the gene sets regulated by C/EBPα and β-catenin-TCF4 may suggest some extent of redundancy or even cooperation between those 2 transcriptional regulators.
Overall, these data establish a role for the canonical Wnt signaling pathway in steady-state granulopoiesis, albeit with relatively modest effects. However, given the vital importance of adequate neutrophil numbers, especially during times of hematopoietic challenge such as systemic infection, the effect of loss of β-catenin-TCF4–mediated transcription on steady-state granulopoiesis may underestimate the true clinical relevance. Therefore, Danek et al test the functional consequences of impaired β-catenin-TCF4–mediated transcription in various models of emergency hematopoiesis including lipopolysaccharide (LPS) stimulation mimicking systemic gram-negative bacterial infection, Candida albicans infection, and 5-fluorouracil (5-FU)–induced BM cell depletion and cytopenia. Collectively, these results reveal not only an impaired emergency granulopoietic response but also a significant survival disadvantage in mice with impaired canonical Wnt signaling challenged with C albicans or 5-FU, thereby demonstrating the clinical significance of intact β-catenin-TCF4 transcriptional activity.
The authors also provide preliminary evidence for the conservation of the role of the canonical Wnt signaling pathway in granulopoiesis across species. Genetic and pharmacological inhibition or stimulation of the Wnt pathway in human CD34+ HSPCs leads to a decreased or increased propensity for granulocytic differentiation, respectively.
The study by Danek et al adds important novel insights into the regulation of steady-state and emergency granulopoiesis. However, a number of important questions remain unanswered. An obvious question is the upstream regulation of Wnt signaling. Which of the more than a dozen Wnt ligands stimulate granulocytic differentiation from HSPCs? Which cell types secrete these Wnt ligands, and where are they located? Do these ligands increase during emergency conditions, and if so, how is this secretion being regulated by, for instance, an inflammatory insult such as infection?
Finally, it might be tempting to explore the use of pharmacological stimulation of canonical Wnt signaling8 to boost granulopoiesis in clinical settings of neutropenia such as congenital neutropenia, inherited BM failure syndromes, or the myelodysplastic syndromes. However, it will be important to gain further insight into the role of canonical Wnt signaling in the regulation of normal HSCs and their malignant counterparts in myeloid neoplasms9 to prevent unintended adverse effects.
Conflict-of-interest disclosure: S.B. declares no competing financial interests.
