

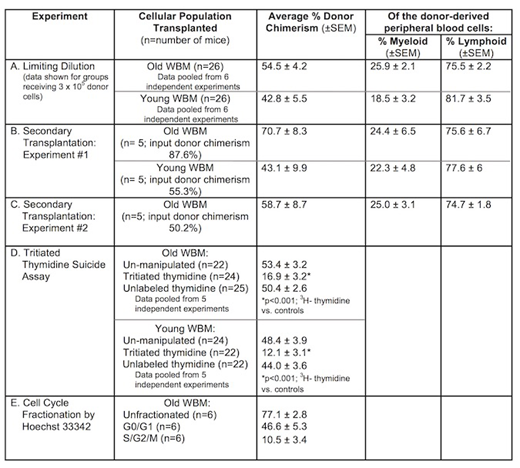
Aging is accompanied by loss of tissue homeostasis, deterioration in organ function and an increased incidence of disease. Age-related changes in the adult stem/progenitor compartment likely drive much of this aging phenotype. For the hematopoietic system, it has been shown that the number of hematopoietic stem cells (HSCs) increases with age, but their renewal potential decreases and they exhibit aberrant lineage production with myeloid skewing. These age-related findings have been based primarily on studies of highly purified, immunophenotypically defined HSCs. However, our data indicate that young murine WBM contains a large population of cycling HSCs not represented within the purified HSC population. The goal of this work was to determine the effects of aging on this total population of stem cells in un-separated WBM. We first performed limiting dilution competitive bone marrow transplantation assays to determine the relative frequency of HSCs in old and young murine WBM. We injected serial dilutions of WBM from either old (24 month old) or young (6-8week old) donor C57BL/6 (CD45.2) mice into lethally irradiated young B6.SJL (CD45.1) hosts. Each mouse received 3x105 competitor WBM cells derived from young B6.SJL mice. At 6 months post transplant, we measured peripheral blood chimerism and lineage composition. According to the Poisson distribution, the estimated frequency of stem cells (with positive engraftment defined as ≥1% donor chimerism) was 1 in 9.8 x 103 for old marrow and 1 in 40 x 103 for young marrow, an approximately 4-fold increase in functional stem cells in old marrow, similar to reports in the literature. Interestingly, in contrast to what has been observed with old purified HSCs, the lineage analysis of donor-derived peripheral blood from aged WBM showed minimal to no myeloid skewing (Table, Row A). To test stem cell function in secondary transplantation, lethally irradiated young recipient mice were initially transplanted with old or young donor WBM mixed with equal numbers of young competitor WBM. At 6 months post-transplant, WBM was harvested and serially transplanted into lethally irradiated secondary recipient young mice. Peripheral blood chimerism was analyzed at 6 months. Similar to young WBM, the stem cells within old WBM retained robust stem cell capacity in serial transplantation, and aged WBM continued to show little to no myeloid skewing (Table, Row B, C). Finally, using tritiated thymidine (3H-thymidine) suicide and Hoechst 33342 staining, we determined the cell cycle status of the aged HSCs in WBM. WBM harvested from old or young donor mice was exposed to 3H-thymidine in vitro for 30 minutes, during which time any cell traversing S-phase would be selectively killed. Cells were then mixed with equal numbers of un-manipulated young competitor WBM and transplanted into lethally irradiated young host mice. Controls included donor WBM un-manipulated or incubated with unlabeled thymidine. We found a significant reduction in engraftment capacity in old WBM post 3H- thymidine incubation (Table, Row D), suggesting the presence of cycling HSCs. Utilizing Hoechst 33342, we isolated G0/G1 and S/G2/M cell cycle-specific fractions of old WBM by FACS and injected each population with equal numbers of young competitor WBM into lethally irradiated young recipient mice. At 6 months post transplant, aged WBM cells in S/G2/M contributed to peripheral blood chimerism, albeit to a lesser extent than cells in G0/G1 (Table, Row E). We previously published that, although purified HSCs in young marrow are quiescent, cells in S/G2/M and in G0/G1 from young un-separated WBM had equal engraftment capacity. Thus, it appears that this cycling stem cell population is still present in old marrow but declines with age. In conclusion, in contrast to purified HSCs, functional engraftment capacity in un-separated WBM does not seem to diminish with age, shows minimal to no myeloid skewing, and is due, in part, to cycling stem cells. These data support the existence of a population of HSCs in WBM distinct from conventionally defined purified HSCs, with seemingly differential susceptibility to aging with respect to self-renewal and lineage fate decisions. Thus, we propose that approaches to reverse aging and define mechanisms underlying age-related hematologic diseases may need to be broadened to incorporate a more heterogeneous HSC compartment not fully represented by the purified HSCs.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal