

Abstract
The ability of human γδ T cells to develop immunologic memory is still a matter of debate. We previously demonstrated the involvement of Vδ2− γδ T lymphocytes in the response of immunosuppressed organ recipients to cytomegalovirus (CMV). Here, we demonstrate their ability to mount an adaptive immune response to CMV in immunocompetent subjects. Vδ2− γδ T-cell peripheral blood numbers, repertoire restriction, and cytotoxicity against CMV-infected fibroblasts were markedly increased in CMV-seropositive, compared with CMV-seronegative, healthy persons. Whereas Vδ2− γδ T cells were found as naive cells in CMV− patients, they virtually all exhibited the cytotoxic effector/memory phenotype in CMV+ patients, which is also observed in transplanted patients challenged with CMV. This long-term complete remodeling of the Vδ2− γδ T-cell population by CMV predicts their ability to exhibit an adaptive anti-CMV immune response. Consistent with this, we observed that the secondary response to CMV was associated with a faster γδ T-cell expansion and a better resolution of infection than the primary response. In conclusion, the increased level of effector-memory Vδ2− γδ T cells in the peripheral blood is a specific signature of an adaptive immune response to CMV infection of both immunocompetent and immunosuppressed patients.
Introduction
Human cytomegalovirus (CMV) is a widespread β-herpesvirus that establishes a lifelong viral persistence without detectable symptoms in immunocompetent patients but with life-threatening consequences in immunologically immature or compromised patients. Many studies have been reported that support an important role for adaptive T lymphocytes in the control of CMV infection.1-4 In addition, we have demonstrated that the γδ T-cell subpopulation contributes to the anti-CMV immune responses.5,6 These unconventional T cells are generally considered to be intermediates between innate and adaptive immunity because of their rapid and massive responses to very diverse immune challenges.
Compelling data exist that demonstrate the importance of γδ T cells in various microbial infections in humans. They exhibit in vitro reactivity against cells infected by viruses, bacteria, or parasites and are selectively expanded in the peripheral blood of infected patients.7-10 The majority of human circulating γδ T cells express a T-cell receptor (TCR) encoded by the Vγ9 and Vδ2 gene segments. These cells are activated after interaction with nonpeptidic phosphorylated compounds, collectively called phosphoantigens, which are metabolic intermediates of the isoprenoid biosynthetic pathway.11,12 These phosphoantigens are expressed by a variety of bacteria and parasites and are also present in some tumors. The other γδ T cells are known as Vδ2− γδ T cells, are largely located in mucosal epithelia and in the spleen, and represent approximately 20% of all circulating γδ T cells. These cells predominantly express a TCR containing the Vδ1 region. Their repertoire in the peripheral blood of most healthy adults is restricted, whereas it is polyclonal in the thymus and in cord blood.13-15 This observation suggests that unknown environmental factors encountered during adult life are selectively amplifying the Vδ1 T lymphocytes subpopulation. The ligand specificity of Vδ1 γδ T cells has been much less explored than that of Vγ9Vδ2 T cells, even though some in vitro isolated Vδ1 γδ T-cell lines or clones have been reported to be activated by CD1c16 or by MHC class I–related chain A or B.17
Although γδ T cells express receptors that are encoded by somatically rearranged genes, they are generally considered to be innate-like effectors unable to establish antigen-specific memory. Nevertheless, adaptive immune responses mediated by Vγ9Vδ2 T cells have been shown to occur in macaques infected with live mycobacteria. The adaptive response to a secondary challenge had shorter kinetics and a higher magnitude Vγ9Vδ2 T-cell response.18 In contrast, injection of an agonist synthetic phosphoantigen leads to γδ T-cell exhaustion in cynomolgus monkeys.19 In humans, phosphoantigen-induced in vitro proliferation of Vγ9Vδ2 T cells is increased by prior Mycobacterium bovis Bacillus Calmette-Guerin vaccination.20,21 This result suggests that human memory Vγ9Vδ2 T cells exist, but in vivo support for this conclusion has not been reported. To our knowledge, the ability of Vδ2− γδ T cells to develop a memory response has never been addressed.
Tissue localization of Vδ2− γδ T cells in physiologic conditions limits their study in humans. However, Vδ2− γδ T cells, but not Vγ9Vδ2 T cells, can persistently expand in the peripheral blood of organ recipients with CMV infection, suggesting that they contribute to the antiviral immune response in this immunocompromised context.5 This suggestion has been confirmed by the demonstration that Vδ2− γδ T-cell expansion is concomitant with the resolution of the viremia22 and that Vδ2− γδ T-cell clones and lines isolated from the patients kill CMV-infected cells and limit viral replication in vitro.6 The immunosuppressed status of transplant patients might be related to this particular expansion and involvement of Vδ2− γδ T cells during the host response against CMV. In this line, similar Vδ2− γδ T-cell expansions associated with CMV infections have been reported in immunodeficient children bearing a hypomorphic mutation in the Recombination Activating Gene-1 gene.23,24 Thus, the role of immunosuppression in the response of Vδ2− γδ T cells to CMV remains unresolved, and studies in immunocompetent patients are needed.
CMV infection in immunocompetent patients is asymptomatic, thus preventing studies on acute primary infections. Therefore, in the present study, we compared the phenotype and the functions of γδ T-cell subsets present in the peripheral blood of healthy immunocompetent patients who had been exposed to CMV and were seropositive, to those who were CMV-seronegative. Strikingly, by selecting random cohorts of blood donors based only on their CMV serologic status, we found major differences of Vδ2− γδ T-cell percentages, repertoire, anti-CMV effector functions, and phenotype. All these features are highly suggestive of a long-term selection of a memory-like γδ T-cell subset. Accordingly, we also showed that previous contact with CMV induced a rapid recall expansion of Vδ2− γδ T cells, which coincided with better infection resolution when CMV was reactivated in transplanted patients. These hallmarks of a secondary response represent the first in vivo demonstration of a human γδ T-cell adaptive antiviral immune response. This study also highlights a very specific relationship between the highly prevalent virus CMV and a still enigmatic subset of human γδ T cells.
Methods
Blood samples from healthy persons
Blood leftovers were obtained from anonymous healthy donors enrolled in the local blood bank cytapheresis program. Samples were subjected to determinations of herpes simplex virus (HSV), varicella-zoster virus (VZV), Epstein-Barr virus (EBV), and CMV serologies using specific enzyme-linked immunosorbent assay kits (Ortho, Eurogentec, or Enzygnost anti-CMV/IgG and IgM; Dade Behring, Deerfield, IL). CMV-seronegative (n = 80) and CMV-seropositive (n = 106) donor groups had comparable mean ages (mean [range] = 40 [19-60] years vs 41 [19-59] years, respectively, P > .05) and sex ratios (male/female = 1.5 vs 0.83, respectively, P > .05).
Renal transplant patients
CHU Bordeaux's ethics committee for the protection of patients involved in biomedical research has approved the study. After giving their written informed consent in accordance with the Declaration of Helsinki, 46 renal transplant patients receiving a standard immunosuppressive regimen (corticosteroids, anticalcineurins, and antimitotics, with or without antithymocyte immunoglobulins) and experiencing CMV infection were analyzed for the delay between CMV infection onset and γδ T-cell expansion in the peripheral blood. The duration of infection was also recorded. Inclusion in the study was based on CMV infection after transplantation and the CMV serology status of the donor and the recipient at the time of the graft. Only D+/R− (n = 25) and D−/R+ (n = 20) patients were studied to compare primary CMV infections to CMV reactivations, respectively. CMV infection was diagnosed based on detection of CMV pp65 antigen (> 1 cell per 2 × 105 leukocytes) in peripheral blood leukocytes (CINAkit; Argene-Biosoft, Varhiles, France).
Flow cytometric analysis
Monoclonal antibodies (mAbs) directed against the following molecules were purchased from Beckman Coulter (Marseille, France): TCR-Cδ, TCR-Vδ1, TCR-Vδ2, TCR-Vδ3, NKG2A, NKp80, CD94, CD57, CD158b/j, CD158a/h, CD158i, and CD158e. mAbs directed against CD3, CD27, CD28, CD45RA, CD62L, perforin, and granzyme B were from BD Biosciences (San Jose, CA), and the anti-NKG2C mAb was from R&D Systems (Minneapolis, MN). Staining was performed on whole blood as previously described and analyzed on a FACScalibur Cytometer or a FACScanto Cytometer (BD Biosciences).5 At least 5000 T lymphocytes were analyzed for 3-color staining and 100 000 T lymphocytes were analyzed for 6-color staining.
Analysis of TCR CDR3-length distribution by Immunoscope
Donor peripheral blood mononuclear cells (PBMCs; 5 × 106) were used to prepare total RNA in TRIzol reagent (Invitrogen, Cergy-Pontoise, France). Total RNA was reverse-transcribed to cDNA with oligo-dT primers and MMLV-reverse transcriptase according to the manufacturer's instructions (Promega, Madison, WI). cDNAs were then amplified with Vδ1 (GCCTTAACCATTTCAGCC), or Vδ2 (TACCGAGAAAAGGACATCTATGGC) and Cδ (GTCGTGTTGAACTGAACATGTCACTG) primers. Forty amplification cycles were performed followed by an additional completion step of 1 hour at 72°C. Aliquots (2 μL) of the polymerase chain reaction products were reacted in a 2-cycle run-off reaction supplemented with the fluorescent CδFAM primer (5′-ACGGATGGTTTGGTATGAGGCTGA-3′). Each labeled product was then loaded onto a 4.5% acrylamide sequencing gel for separation by size. The fluorescence intensity was measured with a DNA sequencer 373A and analyzed with Genescan Software (Applied Biosystems, Foster City, CA). To quantify the deviation of sample T-cell repertoires from controls as measured by Immunoscope, we introduced an index of oligoclonality as described previously.5 This index is defined as follows:
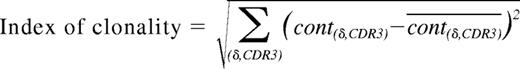
where cont(δ,CDR3) is the contribution of the (δ,CDR3) combination to the donor repertoire and cont(δ,CDR3) is the contribution of the (δ,CDR3) combination to the average repertoire.
Generation of polyclonal γδ T-cell lines
Vδ2− γδ T-cell subpopulations were sorted from blood donor PBMCs using a combination of anti-Vδ1 mAb, anti-Vδ3 mAb, and magnetic beads coated with sheep anti–mouse Ig (Invitrogen). Bead-adherent cells were further expanded in culture with RPMI medium supplemented with 10% human serum, 1000 U/mL of rIL-2 (Chiron, Suresne, France), 1 μg/mL of PHA-L (Sigma-Aldrich, St Louis, MO), and irradiated allogeneic PBMCs (35 Gy). After 1 month of culture, T-cell lines were phenotyped by flow cytometry and used in the CD107 cytotoxicity assay.
CD107 cytotoxicity assay
Subconfluent monolayers of primary foreskin fibroblasts (FSFs) were incubated with a CMV suspension (TB40/E strain) at a multiplicity of infection of 1 and cultured for 5 days at 37°C. Infected cell layers were washed before being used for coculture experiments with γδ T-cell clones. The cytotoxic potential of γδ T-cell lines was measured by the flow cytometric CD107a assay as previously described.25 Briefly, γδ T cells were cocultured with infected or uninfected FSFs for 6 hours in the presence of anti-CD107a mAb (H4A3, BD Biosciences). Brefeldin A was added in the coculture for the last 5 hours. Then cells were harvested, surface stained with anti-Cδ and anti-CD3 mAbs, and analyzed by flow cytometry.
Statistical analysis
Data are expressed as median (range) or mean plus or minus SD. The statistical differences between groups of CMV− and CMV+ donors were tested with the unpaired Mann-Whitney U test using Statview software (Abacus Concepts, Berkeley, CA). Differences were considered to be significant when P was less than .05.
Results
The percentage of circulating Vδ1 γδ T cells is higher in CMV-seropositive (CMV+) donors than CMV-seronegative (CMV−) donors
We previously demonstrated that Vδ2− γδ T cells participate in responses to CMV in immunosuppressed allograft recipients. To evaluate whether Vδ2− γδ T cells are also involved in this response in immunocompetent patients, we measured and compared the percentages of γδ T-cell subsets expressing the 3 main Vδ chains (Vδ1, Vδ2, and Vδ3) in the peripheral blood of CMV+ (n = 80) and CMV− (n = 106) healthy blood donors. As expected, the total γδ T-cell population was mainly composed of Vδ2 cells, which had the same mean percentage in both groups of donors (Figure 1A, Figure S1, available on the Blood website; see the Supplemental Materials link at the top of the online article). However, the percentages of Vδ1 cells were significantly higher in CMV+ than in CMV− patients (P < .001, Figure 1A), although far lower than those observed in transplant patients (∼10%).26 In all groups, Vδ3 T cells were barely detectable using flow cytometry, and it was therefore difficult to make comparisons between groups for this subset. Consistent with previous results, a statistically significant increase of CD8highCD57+ αβ T cells was observed in CMV+ patients (P = .005).27 Because these phenotypic tests were performed on blood samples collected over a period of several months, 5 subjects from this cohort were blindly analyzed twice at distinct blood collection times. They were considered only once within the cohort of 186 donors for the statistical analysis. Notably, 1 subject who seroconverted in the course of the study showed an increase in Vδ1+ T-cell levels between the first and the second blood test (Figure 1B). In conclusion, there is an increased number of circulating Vδ1 T cells in CMV+ immunocompetent patients. This is probably the signature of a prior γδ T-cell selection in the course of CMV infection.

Higher percentages of Vδ1+ γδ T cells in CMV+ vs CMV− healthy subjects. The percentage of γδ T cells in the freshly collected peripheral blood of 80 CMV− and 106 CMV+ blood donors was determined by flow cytometry. Representative raw data are presented in Figure S1. (A) The medians, 25th and 75th percentiles (bottom and top of histograms), and 10th and 90th percentiles (bottom and top bars) of γδ, Vδ2+, Vδ1+, Vδ3+ T cells in the T-lymphocyte population, and CD8highCD57+ cells in the αβ T-cell population. (B) In the course of the study, blind phenotyping of Vδ1+ and Vδ3+ γδ T-cell populations was performed twice for 5 subjects with an interval of several weeks. ○ represent seronegative subjects;  , seropositive subjects; □ and
, seropositive subjects; □ and  , subject presenting a CMV seroconversion between first and second sample.
, subject presenting a CMV seroconversion between first and second sample.
Higher percentages of Vδ1+ γδ T cells in CMV+ vs CMV− healthy subjects. The percentage of γδ T cells in the freshly collected peripheral blood of 80 CMV− and 106 CMV+ blood donors was determined by flow cytometry. Representative raw data are presented in Figure S1. (A) The medians, 25th and 75th percentiles (bottom and top of histograms), and 10th and 90th percentiles (bottom and top bars) of γδ, Vδ2+, Vδ1+, Vδ3+ T cells in the T-lymphocyte population, and CD8highCD57+ cells in the αβ T-cell population. (B) In the course of the study, blind phenotyping of Vδ1+ and Vδ3+ γδ T-cell populations was performed twice for 5 subjects with an interval of several weeks. ○ represent seronegative subjects; , seropositive subjects; □ and , subject presenting a CMV seroconversion between first and second sample.
Influence of CMV infection on the repertoire of Vδ1 γδ T cells
If Vδ1 γδ T cells are selectively expanded by CMV challenge, they should display a restricted repertoire. Several studies have reported that restricted repertoire of Vδ1 T cells can be found in some healthy persons, but the reason for this is unknown.13-15,28 To address this issue, complementary determining region 3 (CDR3) size distributions of TCR Vδ1 chains expressed by PBMCs were analyzed by the Immunoscope technique in 10 CMV+ and 10 CMV− healthy blood donors. As shown in Figure 2, the repertoire of Vδ1 T cells, but not that of Vδ2 T cells, was more restricted in seropositive patients than in seronegative patients. We next used a quantitative index of clonality, which measures the deviation between the repertoire of a given donor and an average repertoire computed as the arithmetic mean of the repertoire of all the donors.5 Accordingly, we observed a correlation (P < .003) between the indices of clonality of Vδ1 T cells, but not that of Vδ2 T cells, and the CMV serology status of the patient (Figure 2A,B left panels). This result demonstrated once again that the Vδ2− γδ T-cell involvement in the anti-CMV immune response is unique.

Oligoclonality of Vδ1, but not Vδ2, repertoire was more pronounced in CMV+ compared with CMV− subjects. Profiles of Cδ-Vδ1 (A) and Cδ-Vδ2 (B) CDR3 length distribution were obtained by Immunoscope analysis of the PBMCs of 10 CMV− and 10 CMV+ donors. (Left panels) Fluorescence intensity is represented in arbitrary units as a function of the size of single-stranded DNA fragments. (Right panels) The index of clonality for each patient studied was calculated as indicated in “Analysis of TCR CDR3-length distribution by Immunoscope” and plotted as a function of the donor CMV (top panels) or HSV (bottom panels) serology status. HSV serology could not be determined for one CMV-seropositive donor, explaining why only 19 donors are plotted in the bottom panels. Means are indicated by a short line.
Oligoclonality of Vδ1, but not Vδ2, repertoire was more pronounced in CMV+ compared with CMV− subjects. Profiles of Cδ-Vδ1 (A) and Cδ-Vδ2 (B) CDR3 length distribution were obtained by Immunoscope analysis of the PBMCs of 10 CMV− and 10 CMV+ donors. (Left panels) Fluorescence intensity is represented in arbitrary units as a function of the size of single-stranded DNA fragments. (Right panels) The index of clonality for each patient studied was calculated as indicated in “Analysis of TCR CDR3-length distribution by Immunoscope” and plotted as a function of the donor CMV (top panels) or HSV (bottom panels) serology status. HSV serology could not be determined for one CMV-seropositive donor, explaining why only 19 donors are plotted in the bottom panels. Means are indicated by a short line.
We then tested whether CMV seropositivity was linked to other herpesvirus seropositivity, which might bias the aforementioned results. All 20 donors were seropositive for VZV and only one was negative for EBV, indicating that these viruses could not be held responsible for the observed restriction of the Vδ1 repertoire in some subjects. Nine patients were seropositive for HSV-1, among whom 5 were also CMV+ and 4 were CMV−. Ten patients were HSV-seronegative, among whom 4 were CMV+ and 6 were CMV−. These results indicate that there was not a correlation between CMV and HSV seropositivity. No statistical correlation between Vδ1 or Vδ2 repertoire restriction and HSV seropositivity could be observed (Figure 2 bottom right). In summary, CMV infection was linked to the Vδ2− γδ T-cell repertoire in the competent immune system.
Amplification of Vδ2− γδ T cells reactive against CMV-infected cells in CMV+ subjects
To determine whether these expanded Vδ2− γδ T cells from CMV+ subjects were reactive against CMV, Vδ1+ and/or Vδ3+ γδ T cells were sorted from the donor PBMCs. Four CMV+ donors with more than 1% Vδ2− γδ T cells, and 6 CMV− donors with less than 0.5% of Vδ2− γδ T cells were analyzed. The sorted Vδ1+ and Vδ3+ γδ T cells were polyclonally expanded with PHA-L, IL-2, and allogeneic PBMCs. This in vitro polyclonal activation has been previously shown to maintain the initial in vivo repertoire of T cells.29 When incubated for 6 hours with CMV-infected FSF monolayers, a significantly higher proportion of Vδ2− γδ T cells from CMV+ donors expressed membrane CD107a, a sensitive marker of cytotoxic activity, in comparison to cells from CMV− donors (Figures 3, S2). As a negative control, uninfected FSFs induced a minimal membrane expression of CD107a on T cells. The mean percentages plus or minus SD of CMV-reactive Vδ2− γδ T cells (the percentage of CD107a+ cells on CMV-infected FSFs minus the percentage of CD107a+ cells on noninfected FSFs) were 13.7% plus or minus 8.8% for the 4 CMV+ donors and 2.7% plus or minus 2.4% for the 6 CMV− donors (P = .01). These results indicate that at least some of the expanded Vδ2− γδ T cells in CMV+ patients were specific for CMV-induced antigenic determinants expressed by infected fibroblasts.

Anti-CMV reactivity of polyclonal Vδ2− γδ T-cell lines isolated from CMV seronegative or seropositive blood donors. Whole Vδ2− γδ T-cell lines generated by polyclonal activation were incubated on monolayers of CMV-infected or uninfected primary fibroblasts in the presence of an anti-CD107a mAb. After 6 hours, T cells were stained with an anti-Cδ mAb and analyzed by flow cytometry; representative raw data are presented in Figure S2. Shown here are the percentages of CD107a+ cells in γδ T cells for each studied donor.
Anti-CMV reactivity of polyclonal Vδ2− γδ T-cell lines isolated from CMV seronegative or seropositive blood donors. Whole Vδ2− γδ T-cell lines generated by polyclonal activation were incubated on monolayers of CMV-infected or uninfected primary fibroblasts in the presence of an anti-CD107a mAb. After 6 hours, T cells were stained with an anti-Cδ mAb and analyzed by flow cytometry; representative raw data are presented in Figure S2. Shown here are the percentages of CD107a+ cells in γδ T cells for each studied donor.
Opposite effector versus naive phenotype of Vδ2− γδ T cells in CMV+ and CMV− donors, respectively
To test whether these expanded Vδ2− γδ T cells had memory activity, we analyzed the phenotype of freshly collected PBMCs. Analysis of CD27, CD28, CD45RA, and CD62L expression by Vδ2− γδ T cells revealed strikingly different profiles for CMV− and CMV+ donors (Figures 4A, S3). Vδ2− γδ T cells expressed mainly a CD45RA+ CD27+ CD28+/− CD62L+ naive phenotype in CMV− donors. In sharp contrast, they were predominantly CD45RA+ CD27− CD28− CD62L− effector/memory (TEMRA) cells in CMV+ donors. As shown in Figure 4, the variability of the expression of CD45RA/CD27 was low between patients within the tested cohort. The mean percentage of TEMRA cells in the Vδ2− γδ T-cell population was 13.7% plus or minus 7.4% in CMV− donors (n = 9) and 68.4% plus or minus 15.7% in CMV+ donors (n = 9; P < .001). Vδ1 and Vδ3 T cells (which comprise > 99% of Vδ2− γδ T cells) were analyzed separately in some CMV+ donors. As expected, Vδ1 were the dominant subpopulation of Vδ2− γδ T cells and were mainly TEMRA cells in all but one donor (Figure S4). Vδ3 T cells had a TEMRA phenotype in the donors exhibiting the highest percentages of Vδ3 T cells (Figure S4, donors 1-5), and a naive phenotype in the rest (data not shown). The Vδ2+ T cells clearly exhibited the previously reported CD45RA− CD27+ CD28+/− CD62L− central memory phenotype30-32 in both CMV− and CMV+ donors. The TEMRA Vδ2+ T cell minor subset showed a slight but significant increase in CMV+ donors (P = .04). TEMRA Vδ2− γδ T cells in CMV+ donors expressed high amounts of perforin and granzyme B, in contrast to the naive Vδ2− γδ T cells of CMV− donors (Figure S4). A majority of these TEMRA Vδ2− γδ T cells from CMV+ donors expressed NK cell receptors, such as the killer Ig-like receptors (CD158) or NKp80, and the senescence marker CD57 (Figures 4B, S4). A subset of these cells also expressed CD94 in association with NKG2C. In contrast, naive Vδ2− γδ T cells from CMV− donors were predominantly devoid of CD57, NKp80, and CD158 expression.

The expanded Vδ2− γδ T cells in CMV-experienced donors are mainly TEMRA cells. Freshly collected blood from CMV− or CMV+ blood donors was stained for 6-color flow cytometric analysis with mAbs directed against CD3, γδ TCR, Vδ2 and activation markers or NKR. Raw data are presented in Figure S3. (A) Percentages of Vδ2+ or Vδ2− γδ T cells positive for CD27 and/or CD45RA in 9 CMV− ( ) and 9 CMV+ donors (●). The short line represents the mean percentage for each cohort. (B) Mean (± SD) percentages of positive cells (for CD57, CD158a/h, b/j, i, e, CD94, NKG2A, NKG2C, or NKp80) or negative cells (for CD27) among Vδ2− γδ T cells from CMV− or CMV+ blood donors.
) and 9 CMV+ donors (●). The short line represents the mean percentage for each cohort. (B) Mean (± SD) percentages of positive cells (for CD57, CD158a/h, b/j, i, e, CD94, NKG2A, NKG2C, or NKp80) or negative cells (for CD27) among Vδ2− γδ T cells from CMV− or CMV+ blood donors.
The expanded Vδ2− γδ T cells in CMV-experienced donors are mainly TEMRA cells. Freshly collected blood from CMV− or CMV+ blood donors was stained for 6-color flow cytometric analysis with mAbs directed against CD3, γδ TCR, Vδ2 and activation markers or NKR. Raw data are presented in Figure S3. (A) Percentages of Vδ2+ or Vδ2− γδ T cells positive for CD27 and/or CD45RA in 9 CMV− () and 9 CMV+ donors (●). The short line represents the mean percentage for each cohort. (B) Mean (± SD) percentages of positive cells (for CD57, CD158a/h, b/j, i, e, CD94, NKG2A, NKG2C, or NKp80) or negative cells (for CD27) among Vδ2− γδ T cells from CMV− or CMV+ blood donors.
Adaptive immune response of Vδ2− γδ T cells during CMV infection in organ transplantation patients
We next examined whether the expanded Vδ1+ T cells in CMV+ patients act as memory cells and are able to manifest a rapid recall response. To answer this question, the kinetics of γδ T-cell expansion was studied in CMV+ patients transplanted with CMV− renal allografts (donor−/recipient+ [D−/R+], n = 20) and who underwent secondary viral challenge because of CMV reactivation during immunosuppressive treatment. They were compared with CMV− recipients transplanted with a CMV+ graft (D+/R−, n = 25), who were experiencing CMV primary infections. As previously described,26 all patients had an expansion of Vδ2− γδ T cells in their peripheral blood. However, a more rapid expansion of Vδ2− γδ T cells was observed in patients with CMV reactivations (mean, 17 days; range, −21 to 126 days after onset of CMV infection) vs those with primary infections (mean, 66 days; range, 27 to 240 days, P < .005; Figure 5A). As previously reported, this faster response was associated with a faster resolution of CMV infection because D−/R+ patients displayed a positive CMV− antigenemia for 4.5 (range, 1-9) weeks versus 10 (range, 2-20) weeks for D+/R− patients (P < .001; Figure 5B). The phenotype of Vδ2− γδ T cells was analyzed longitudinally before and after CMV infection in 3 D+/R− and 3 D−/R+ transplant patients. As shown in Figure 5C, D+/R− patients exhibited much lower percentages of TEMRA Vδ2− γδ T cells in comparison to D−/R+ patients, a distribution very similar to that observed in healthy donors (Figure 4). During the course of CMV infection, an increase of TEMRA Vδ2− γδ T-cell percentages appeared in the D+/R− patients, suggesting that CMV drives remodeling of the naive cell phenotype toward an effector/memory profile. As expected, the high percentage of TEMRA Vδ2− γδ T cells remained stable in D−/R+ patients during CMV infection (Figure 5C). Therefore, we propose that CMV infection led to an expansion of a pool of memory-like effector Vδ2− γδ T cells which were able to rapidly respond to a second encounter with CMV.

A faster expansion of γδ T cells correlates with a faster infection resolution during CMV reactivation compared with primary infection. Forty-six renal allotransplantation patients, selected because they developed CMV infections, were divided according to the CMV serology of the recipient (R) and the donor (D) at the time of the graft. All patients displayed γδ T-cell expansion in their peripheral blood. (A) The time to γδ T-cell expansion was defined for each patient as the interval between the first day of CMV detection in the peripheral blood (pp65 antigenemia) and the day when γδ T cell levels plateaued.22 (B) The duration of infection was defined as the number of weeks with a positive pp65 antigenemia. The long dashes in panels A and B represent the median value for each cohort. (C) 3 D+/R− and 3 D−/R+ transplant patients who developed a CMV infection were monitored longitudinally for CD45RA and CD27 expression by Vδ2− γδ T cells. Represented are the percentages of CD45RA+CD27− cells in the Vδ2− γδ T-cell population before and after CMV infection.
A faster expansion of γδ T cells correlates with a faster infection resolution during CMV reactivation compared with primary infection. Forty-six renal allotransplantation patients, selected because they developed CMV infections, were divided according to the CMV serology of the recipient (R) and the donor (D) at the time of the graft. All patients displayed γδ T-cell expansion in their peripheral blood. (A) The time to γδ T-cell expansion was defined for each patient as the interval between the first day of CMV detection in the peripheral blood (pp65 antigenemia) and the day when γδ T cell levels plateaued.22 (B) The duration of infection was defined as the number of weeks with a positive pp65 antigenemia. The long dashes in panels A and B represent the median value for each cohort. (C) 3 D+/R− and 3 D−/R+ transplant patients who developed a CMV infection were monitored longitudinally for CD45RA and CD27 expression by Vδ2− γδ T cells. Represented are the percentages of CD45RA+CD27− cells in the Vδ2− γδ T-cell population before and after CMV infection.
Discussion
We have previously described a correlation between CMV infection and γδ T-cell expansion in immunosuppressed transplant patients.26 This association was also documented in immunodeficient children with a hypomorphic Recombinant Activating Gene-1 mutation.23,24 Both situations lead to a deficiency of the adaptive immune system, which raises the possibility that immunosuppression drives γδ T cells to play a surrogate function in the anti-CMV immune response. One aim of the present study was therefore to determine whether an expansion of γδ T cells could be detected in immunocompetent patients who experienced a CMV infection. The γδ T-cell subset that expanded in transplant patients (ie, Vδ2− γδ T cells) was also found to be increased in healthy CMV-seropositive patients compared with CMV-seronegative patients. These cells displayed a restricted repertoire, which can be regarded as the signature of a CMV-driven selection and amplification of specific T cells in vivo. In agreement with this, they comprised an increased proportion of the cells displaying in vitro CMV-specific cytotoxicity. Altogether, these data indicate an involvement of the γδ T cells in a competent immune response to CMV.
Notably, the percentage of Vδ2− γδ T cells in the peripheral T cells of healthy CMV+ patients was much lower than in transplant patients infected with CMV (1.2% vs > 10.0%, respectively).26 In allograft patients, peripheral blood γδ T cells remained elevated for at least 10 years, the time of last follow-up. We can only speculate that this might be the result of the long-term immunosuppressive treatment that continues to affect T-cell homeostasis33 and thus promotes γδ T-cell persistence in the periphery, and/or to low-grade sporadic multiplication of CMV in selected niches. The lower percentage of γδ T cells that is detected in CMV+ healthy persons compared with transplant patients might be explained by a constrained expansion in response to CMV challenge and/or a negative regulation loop that is normally initiated after the resolution of the infection but that was impaired by immunosuppressive drugs. CMV infection is asymptomatic or subclinical in the majority of immunocompetent patients. Thus, the status of acute primary infections remains ethically and technically out of reach, making this question difficult to address directly. Nevertheless, we can reasonably state that the large γδ T-cell expansion that was observed in the immunosuppressed patients is an amplified form of the normal immune response to CMV.
Several previous studies reported on the restricted repertoire of Vδ1 T cells that is commonly found in healthy adults.13-15 No physiologic or environmental cause was proposed to explain this phenomenon. We showed in this study that the restriction of the Vδ1 T-cell repertoire is associated with CMV but not with HSV, VZV, or EBV infection. We have previously reported additional support for this relationship in transplant patients for whom γδ T-cell expansion is not correlated with viral infections (HSV, VZV, EBV, influenza),26 and in vitro where Vδ2− γδ T-cell clones are able to kill CMV-infected fibroblasts but have no effect on HSV- or VZV-infected fibroblasts.6 In the present study, we compared 2 groups of donors distinguished only by their CMV serotype status. Despite all the other pathogens encountered by each individual donor, the majority of Vδ2− γδ T cells were naive in CMV− donors yet were memory effectors in CMV+ donors. This contrasting phenotype emphasizes the striking relationship between this lymphocyte subset and CMV and suggests that all circulating effector/memory Vδ2− γδ T cells are anti-CMV-specific T cells. Such a dramatic skewing of the Vδ2− γδ T-cell phenotype provides a useful and easy tool for the diagnosis of active or latent CMV infection. It could prove to be a sensitive and specific biologic marker in addition to or as an alternative to CMV serology for the detection of CMV infections in patients. However, analysis of large cohorts of donors is required to compare rigorously the specificity and sensitivity of each test.
Given the prevalence of CMV seropositivity worldwide, this virus appears to be a major environmental factor involved in the formation of an adult Vδ2− γδ T-cell repertoire. This is reminiscent of significant impact of CMV on the CD4 and CD8 αβ T-cell repertoire. The proportion of CD4 and CD8 T cells that are specific for CMV in healthy persons previously exposed to this virus has been shown to be strikingly high (up to 40% of memory T cells).34 This frequency exceeds those observed for other common viruses, and CMV can also reduce prevailing levels of immunity to other persistent viruses.35 An increase of CMV-specific CD8 T cells has been demonstrated in elderly donors and is postulated to be result of life-long recurrent reactivations.36 The presence of cells permissive to CMV replication (mainly fibroblasts, epithelial cells, and endothelial cells) in nearly all organs may play an important role in this wide activation of CMV-specific T cells.37 In this context, our results identify Vδ2− γδ T cells as an additional, statistically significant, component of the immune response to CMV infections.
The long-term expansion of Vδ2− γδ T cells in CMV-experienced patients displayed effector/memory T-cell features resembling that of CMV-specific CD4 or CD8 αβ T cells analyzed in previous studies.38,39 This dominant representation of CD45RA+ CD27− cells among virus-specific αβ T cells is a characteristic of CMV infection that is not found with infections of other persistent viruses, such as EBV, HIV, or HCV.38,40 The reason why different chronic viral infections elicit distinct immune commitment is not clear but may relate to the different sites of virus replication. The long-lived effector/memory CD8 T cells poised for immediate response to infection have been shown to preferentially home to nonlymphoid tissue on antigenic challenge.41 Accordingly, Vγ9Vδ2 T cells found in ascites and cerebrospinal fluids of tuberculosis patients predominantly express a TEMRA phenotype.42 Similarly, Vδ2− γδ T cells are found in the gastrointestinal tract, which is the preferential site of CMV replication. This could be a local mechanism for rapid containment of the virus by direct recognition and elimination of infected cells. The TEMRA Vδ2− γδ T cells that we observed in the peripheral blood could be patrolling cells migrating from these sites with the goal of reaching other infected tissues. The local expansion of Vδ2− γδ T cells could have been overrepresented in immunosuppressed patients because of their impaired αβ or regulatory T cells.43
Despite compelling evidence that γδ T cells can protect mice against several types of infection, their ability to develop pathogen-specific immunity has never been established in humans. In macaques, a memory-type response and rapid expansion of Vγ9Vδ2 T cells have been observed on a secondary challenge with Bacillus Calmette-Guerin.18 In contrast to these results, a study performed in cynomolgus monkeys that were treated with a synthetic ligand of Vγ9Vδ2 T cells showed a diminished ability of these cells to expand on repeated challenge with the agonist, suggesting a progressive exhaustion of the response.19 The role of Vδ2− γδ T cells in CMV infection provides an interesting model to follow the response of these cells to primary or secondary challenge with this virus in humans. The demonstration of a long-term expansion of Vδ2− γδ T cells with a restricted repertoire in CMV-seropositive patients, and, more importantly, of their quicker response to a secondary challenge in graft recipients, strongly suggests that they participate in the adaptive response. Furthermore, they share the same effector/memory phenotype as CMV-experienced CD4 and CD8 T cells, supporting the notion that all these T-cell subsets take part in the adaptive immune response. Combined with our previous observations that delayed expansion of γδ T cells correlates with the severity of CMV infection in transplanted patients22 and that Vδ2− γδ T cells can kill CMV-infected cells in vitro,6 the results obtained in this study form a solid basis to propose Vδ2− γδ T cells as an active component of the antiviral response. A direct evidence of a protective role of these lymphocytes against CMV will require studies in nonhuman primate models of CMV infection.
The development of a CMV vaccine has been an important goal for at least 2 decades. It is well known that both humoral and cellular immune responses have crucial roles in controlling CMV infection. However, a better understanding of the complex network built by the different cellular components of the innate and adaptive immune responses is still needed to design a vaccine that is efficient and protective. Thus, the integration of γδ T cells into this network and examination of their potential contributions to immunotherapeutic approaches designed to control or prevent CMV replication deserve further consideration.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
The authors thank the Etablissement Français du Sang d'Aquitaine for blood samples, the nursing staff of the kidney transplant department, J. C. Carron, M. Garcie, and F. Saussais (Laboratoire d'Immunologie, Centre Hospitalier Universitaire de Bordeaux) for skillful routine immunological analyses, B. Tauzin (Université Bordeaux 2) for assistance in the immunoscope analysis, C. Schlundt (CNRS UMR 5164) for technical assistance, C. Sinzger (Institut für Medizinische Virologie und Epidemiologie der Viruserkrankungen, Tübingen, Germany) for the kind gift of the CMV strain TB40/E, C. Pain (Inserm U876, Université Bordeaux 2) for providing FSF, and C. Behr and M. Capone (CNRS UMR 5164) for critical reading of the manuscript.
This work was supported in part by institutional grants from CNRS and the Ministère de l'Enseignement Supérieur et de la Recherche and by grants from the Agence National de la Recherche (ANR-05-JCJC-0129-01), the Fondation pour la Recherche Médicale, the Délégation à la Recherche Clinique d'Aquitaine, and the Conseil Régional d'Aquitaine.
National Institutes of Health
Authorship
Contribution: V.P., D.R., and X.L. performed laboratory research and collected and analyzed data; L.C. and P.M. designed the clinical research methods and analyzed clinical data; I.G. and M.-E.L. designed and collected the samples for the viral analysis; J.-F.M. designed the research approach and wrote the paper; and J.D.-M. designed the research approach, analyzed and interpreted data, and wrote the paper.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Julie Déchanet-Merville, UMR CNRS 5164, Université Bordeaux 2, 146 rue Léo Saignat, F-33076 Bordeaux Cedex, France; e-mail: julie.dechanet@u-bordeaux2.fr.

{kind=link}
This feature is available to Subscribers Only
Sign In or Create an Account Close Modal