TO THE EDITOR:
Hepcidin is a key iron regulatory hormone that downregulates the iron exporter ferroportin to limit iron entry into the circulation from dietary sources and body stores.1 Hepcidin is suppressed by the bone marrow–derived hormone erythroferrone (ERFE) in response to erythropoietin (EPO).2 This action has a physiological role to ensure adequate iron for erythropoiesis in the context of anemia but contributes to iron overload in diseases of ineffective erythropoiesis.3 We recently discovered that ERFE suppresses hepcidin by interfering with bone morphogenetic protein (BMP)-SMAD signaling, hepcidin’s central transcriptional regulatory pathway in the liver.4 This signaling pathway is activated by endothelial cell–derived BMP6 and BMP2 ligands5-7 binding to hepatocyte BMP type I and type II receptors8-10 and the coreceptor hemojuvelin11 to phosphorylate SMAD1/5/8 transcription factors,12 which activate the hepcidin promoter. We showed that ERFE and EPO inhibited SMAD1/5/8 phosphorylation and hepcidin transcription in primary hepatocytes and mice but failed to suppress hepcidin when Smad1 and Smad5 were ablated.4 ERFE was subsequently shown to inhibit hepcidin induction by BMP5, BMP6, and BMP7 (but not BMP2, BMP4, or BMP9) in vitro. It was postulated that ERFE suppresses hepcidin by acting as a ligand trap for BMP5/6/7 subfamily ligands.13 However, a direct interaction between ERFE and BMPs was not shown, and it remains possible that ERFE interacts with BMP receptors that preferentially use BMP5/6/7. Importantly, the detailed mechanism by which ERFE suppresses hepcidin in vivo remains unknown. Here, we examined whether ERFE-mediated hepcidin suppression depends on specific BMP receptors in vitro and in mice, and we determined the binding interactions between ERFE, BMP ligands, and BMP receptors.
Primary hepatocytes isolated from C57BL/6x129S5 background mice were cultured as previously described.4 Cells were then transfected with 80 nM of small interfering RNA (siRNA) targeting BMP receptors or control siRNA (Invitrogen) for 30 hours, and treated with conditioned medium containing 50% (vol/vol) supernatant from control HEK293T cells (CTRL-CM) or HEK293T cells overexpressing mouse ERFE (ERFE-CM)2 for 15 hours. Hep3B cells were transfected with 200 ng of mouse Erfe complementary DNA or empty vector (Origene) for 48 hours and stimulated with 5 ng/mL of BMP ligand for 6 hours. Animal protocols were approved by the Institutional Ethics Committee of the North Rhine-Westphalian Agency for Nature, Environment, and Consumer Protection at Muenster University, Muenster, Germany (permit number Az. 84-02.04.2014.A015, Az.84-02.04.2013.A281). Hepatocyte-specific Alk2 (Alk2fl/fl;Alb-Cre) or Alk3 (Alk3fl/fl;Alb-Cre)–deficient mice and Cre- controls on a C57BL/6 background8 were fed a chow diet (198 parts per million iron). Eight-week-old female mice were given an intraperitoneal injection of EPO (200 U; Janssen) or phosphate-buffered saline (PBS) and were euthanized after 15 hours. RNA isolation and quantitative reverse transcription–polymerase chain reactions were performed and analyzed as described4,10,14 by using the primers shown in supplemental Table 1 (available on the Blood Web site). Transcript levels were normalized to Rpl19 or 18S as internal controls. For immunoprecipitation and pull-down assays, 0.5 to 2 μg of ALK3-Fc, ALK2-Fc, BMP2, BMP6, or BMP2/6 (R&D Systems) were mixed with 0.3 to 3 μL of ERFE-FLAG at 52.9 ng/μL concentrated from serum-free ERFE-CM using a 30 KDa Filter Unit (Ambion) in NETN buffer. The ERFE-FLAG concentration was determined by using an enzyme-linked immunosorbent assay (Intrinsic LifeSciences). Samples were incubated with anti-FLAG affinity gel (MilliporeSigma) or protein A agarose (Thermo Scientific) at 4°C overnight, and eluted with 3× FLAG peptide (MilliporeSigma) or 2× Laemmli buffer before immunoblot using antibodies (supplemental Table 2). Data are presented as box plots with minimum to maximum whiskers or mean ± SEM. Means were compared by using the Student paired t test, one-way analysis of variance with Tukey’s post hoc test, or (for nonparametric data sets) the Mann-Whitney U test using Prism 7 (GraphPad Software). P < .05 was considered significant.
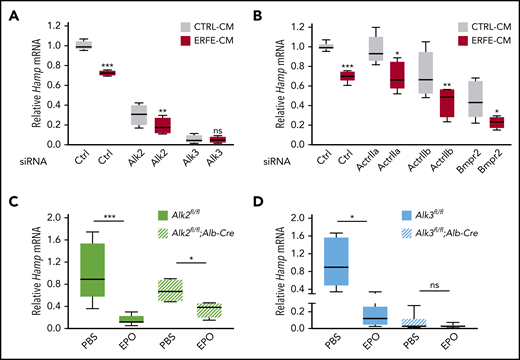
To explore the role of BMP receptors in hepcidin suppression by ERFE, we first performed siRNA knockdown experiments in primary hepatocytes. siRNA knockdown efficiency and specificity were verified (supplemental Figure 1). ERFE suppressed hepcidin messenger RNA (mRNA) in cells transfected with siRNAs targeting each of the three type II receptors and the type I receptor Alk2 but not Alk3 (Figure 1A-B). To determine whether ALK3 is also important for hepcidin suppression by ERFE in vivo, we injected EPO or PBS into mice with or without hepatocyte Alk2 or Alk3 deficiency. EPO similarly induced spleen Erfe mRNA in all mouse groups (supplemental Figure 2) and decreased liver hepcidin mRNA in both Cre- controls (Figure 1C-D). EPO also decreased liver hepcidin in Alk2fl/fl;Alb-Cre mice, albeit to a lesser extent (Figure 1C). However, EPO failed to decrease hepcidin in Alk3fl/fl;Alb-Cre mice (Figure 1D). Although basal hepcidin expression was low in Alk3 siRNA transfected cells and Alk3fl/fl;Alb-Cre mice, hepcidin was further suppressible by serum-free medium (in cells) and a low-iron diet (in mice), showing that hepcidin was not yet at its nadir (supplemental Figure 3). Thus, ALK3 is functionally important for hepcidin suppression by EPO and ERFE, whereas ALK2 plays a minor role.
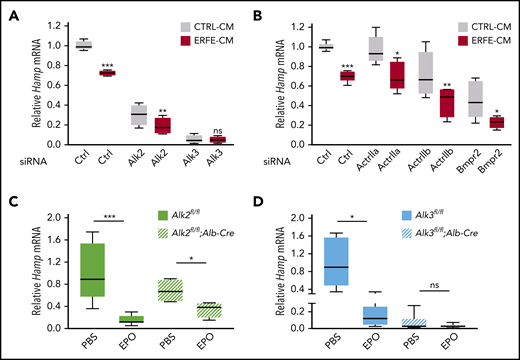
The BMP type I receptor ALK3 is functionally important for hepcidin suppression by ERFE/EPO in primary hepatocytes and in mice. (A-B) Primary hepatocytes were transfected with negative control (Ctrl), Alk2, Alk3, ActrIIa, Actrllb, or Bmpr2 siRNA (80 nM) for 30 hours, followed by treatment with conditioned medium containing 50% (vol/vol) cell supernatant from control HEK293T cells (CTRL-CM) or HEK293T cells overexpressing ERFE (ERFE-CM)2 for 15 hours. Panel A, n = 4 independent experiments; panel B, n = 5 independent experiments. Hepatocyte-specific Alk2-deficient mice (C, Alk2fl/fl;Alb-Cre), hepatocyte-specific Alk3-deficient mice (D, Alk3fl/fl;Alb-Cre) and their respective controls (Alk2fl/fl or Alk3fl/fl; n = 4-8 per group) at 8 weeks of age were given 1 intraperitoneal dose of EPO (200 U per mouse) or PBS, and livers were collected after 15 hours. Relative hepcidin (Hamp) mRNA levels were determined by using quantitative reverse transcription–polymerase chain reaction. Transcripts were normalized to Rpl19 (in cells) or 18S (in mice) as internal controls, and the average of Ctrl cells treated with CTRL-CM or the respective Alk2fl/fl or Alk3fl/fl control mice was set to 1. Data are presented as box plots with minimum to maximum whiskers. *P < .05, **P < .01, ***P < .001 relative to the indicated control by the Student t test or Mann Whitney U test. ns, not significant.
The BMP type I receptor ALK3 is functionally important for hepcidin suppression by ERFE/EPO in primary hepatocytes and in mice. (A-B) Primary hepatocytes were transfected with negative control (Ctrl), Alk2, Alk3, ActrIIa, Actrllb, or Bmpr2 siRNA (80 nM) for 30 hours, followed by treatment with conditioned medium containing 50% (vol/vol) cell supernatant from control HEK293T cells (CTRL-CM) or HEK293T cells overexpressing ERFE (ERFE-CM)2 for 15 hours. Panel A, n = 4 independent experiments; panel B, n = 5 independent experiments. Hepatocyte-specific Alk2-deficient mice (C, Alk2fl/fl;Alb-Cre), hepatocyte-specific Alk3-deficient mice (D, Alk3fl/fl;Alb-Cre) and their respective controls (Alk2fl/fl or Alk3fl/fl; n = 4-8 per group) at 8 weeks of age were given 1 intraperitoneal dose of EPO (200 U per mouse) or PBS, and livers were collected after 15 hours. Relative hepcidin (Hamp) mRNA levels were determined by using quantitative reverse transcription–polymerase chain reaction. Transcripts were normalized to Rpl19 (in cells) or 18S (in mice) as internal controls, and the average of Ctrl cells treated with CTRL-CM or the respective Alk2fl/fl or Alk3fl/fl control mice was set to 1. Data are presented as box plots with minimum to maximum whiskers. *P < .05, **P < .01, ***P < .001 relative to the indicated control by the Student t test or Mann Whitney U test. ns, not significant.
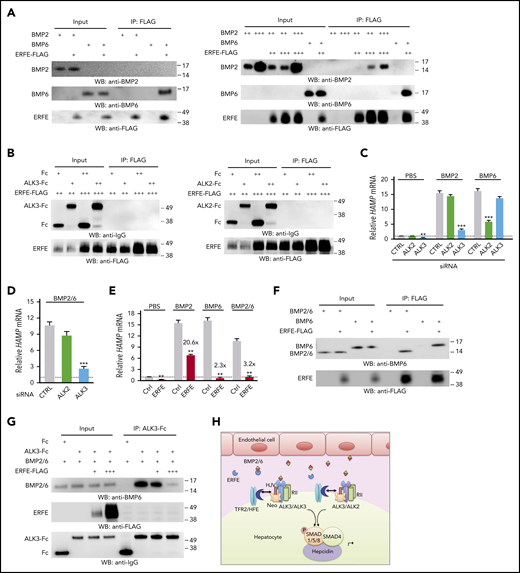
We next used immunoprecipitation/pull-down assays to explore whether ERFE directly binds ALK3 or ALK2. In addition, BMP receptor knockdown experiments were used to examine whether the preferential utilization of different receptor subsets by BMP ligands could account for ERFE’s differential inhibition of BMP5/6/7 but not BMP2/4/9. Finally, we tested whether ERFE directly binds BMP ligands as recently proposed,13 focusing on BMP6 and BMP2, the confirmed physiological regulators of hepcidin in vivo.5-7 ERFE selectively bound BMP6 but required higher concentrations to bind BMP2 (Figure 2A). In contrast, ERFE did not bind ALK3 or ALK2 (Figure 2B; supplemental Figure 4). As positive controls, both BMP2 and BMP6 bound ALK3, and BMP6 bound ALK2 (supplemental Figure 5). However, whereas BMP2 preferentially used ALK3, BMP6 favored ALK2 for hepcidin induction in Hep3B cells (Figure 2C). These data provide the first demonstration of direct binding between ERFE and BMP ligands but not BMP receptors. They also confirm the selectivity of ERFE for binding BMP6 compared with BMP2. However, these data do not explain why EPO failed to suppress hepcidin in Alk3fl/fl;Alb-Cre mice, in which BMP6-ALK2 signaling should remain intact.
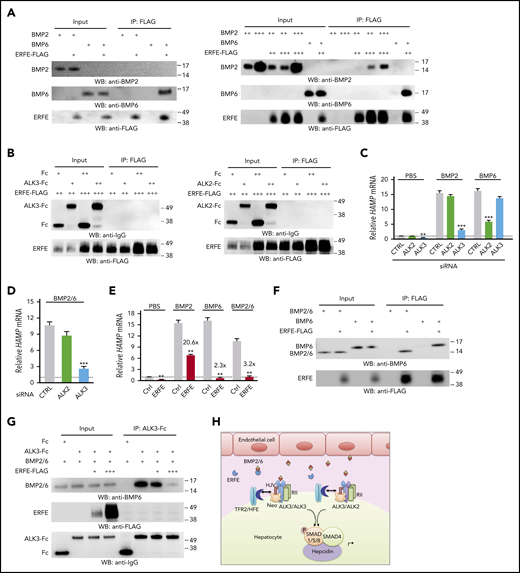
ERFE binds BMP2/6 heterodimer and inhibits hepcidin expression by sequestering BMP2/6 from binding to ALK3. (A) 0.5 μg (+), 2 μg (++), 6 μg (+++) BMP2, or 0.5 μg (+) BMP6 were incubated with 0.3 μL (+), 1 μL (++), or 3 μL (+++) of ERFE-FLAG at 52.9 ng/μl. (B,F-G) 0.5 μg (+) or 2 μg (++) ALK3-Fc, ALK2-Fc, Fc, BMP6, or BMP2/6 were incubated with 0.3 μL (+), 1 μL (++), or 3 μL (+++) of ERFE-FLAG at 52.9 ng/μL. Three percent of the total mixture was saved as input. Samples were immunoprecipitated (IP) with anti-FLAG M2 affinity gel (A-B,F) or protein A agarose (G) in 500 μL of NETN buffer (100 mM NaCl, 1 mM EDTA, 20 mM Tris-HCl, pH 8.0, and 0.5% Nonidet P-40) supplemented with 1× protease inhibitor cocktail (MilliporeSigma P8340) at 4°C overnight. This was followed by elution with 150 μg/mL 3× FLAG peptide in Tris-buffered saline at 4°C for 30 minutes (A-B,F) or 2× Laemmli buffer containing 100 mM of β-mercaptoethanol at 95°C for 5 minutes (G) before sodium dodecyl sulfate–polyacrylamide gel electrophoresis and immunoblot analyses. Experiments were repeated 3 times, with 1 representative blot shown. (C-D) Hep3B cells were transfected with 40 nM of ALK2 or ALK3 siRNA for 30 hours, serum-starved overnight with 1% fetal bovine serum, and treated with PBS or 5 ng/mL of BMP2, BMP6, or BMP2/6 for 6 hours. (E) Hep3B cells were transfected with 200 ng of complementary DNA encoding Erfe or pCMV6-entry empty vector (CTRL) for 48 hours and treated with PBS or 5 ng/mL of BMP2, BMP6, or BMP2/6 in growth medium containing 1% fetal bovine serum for 6 hours. Panels C-E, n = 3 independent experiments. Data are presented as mean ± SEM. **P < .01, ***P < .001 relative to the respective control by the Student t test or one-way analysis of variance with Tukey’s post hoc test. (H) Proposed model depicting ERFE’s mechanism of action: BMP2/6 heterodimeric ligand is secreted by liver endothelial cells and binds to the BMP receptor complex that has been proposed to contain two BMP type II receptors (RII), two type I receptors in the form of ALK3/ALK3 homodimers or ALK2/3 heterodimers, the coreceptor hemojuvelin (HJV), and possibly other interacting proteins, including HFE, transferrin receptor 2 (TFR2), and neogenin (Neo).10,14,17 Activated type I receptors phosphorylate SMAD1/5/8 proteins, which complex with SMAD4 and translocate to the nucleus to induce hepcidin transcription. In the context of erythropoietic drive, secreted ERFE binds to BMP2/6 to prevent BMP2/6 from binding and activating the BMP receptor complex, thereby suppressing hepcidin transcription.
ERFE binds BMP2/6 heterodimer and inhibits hepcidin expression by sequestering BMP2/6 from binding to ALK3. (A) 0.5 μg (+), 2 μg (++), 6 μg (+++) BMP2, or 0.5 μg (+) BMP6 were incubated with 0.3 μL (+), 1 μL (++), or 3 μL (+++) of ERFE-FLAG at 52.9 ng/μl. (B,F-G) 0.5 μg (+) or 2 μg (++) ALK3-Fc, ALK2-Fc, Fc, BMP6, or BMP2/6 were incubated with 0.3 μL (+), 1 μL (++), or 3 μL (+++) of ERFE-FLAG at 52.9 ng/μL. Three percent of the total mixture was saved as input. Samples were immunoprecipitated (IP) with anti-FLAG M2 affinity gel (A-B,F) or protein A agarose (G) in 500 μL of NETN buffer (100 mM NaCl, 1 mM EDTA, 20 mM Tris-HCl, pH 8.0, and 0.5% Nonidet P-40) supplemented with 1× protease inhibitor cocktail (MilliporeSigma P8340) at 4°C overnight. This was followed by elution with 150 μg/mL 3× FLAG peptide in Tris-buffered saline at 4°C for 30 minutes (A-B,F) or 2× Laemmli buffer containing 100 mM of β-mercaptoethanol at 95°C for 5 minutes (G) before sodium dodecyl sulfate–polyacrylamide gel electrophoresis and immunoblot analyses. Experiments were repeated 3 times, with 1 representative blot shown. (C-D) Hep3B cells were transfected with 40 nM of ALK2 or ALK3 siRNA for 30 hours, serum-starved overnight with 1% fetal bovine serum, and treated with PBS or 5 ng/mL of BMP2, BMP6, or BMP2/6 for 6 hours. (E) Hep3B cells were transfected with 200 ng of complementary DNA encoding Erfe or pCMV6-entry empty vector (CTRL) for 48 hours and treated with PBS or 5 ng/mL of BMP2, BMP6, or BMP2/6 in growth medium containing 1% fetal bovine serum for 6 hours. Panels C-E, n = 3 independent experiments. Data are presented as mean ± SEM. **P < .01, ***P < .001 relative to the respective control by the Student t test or one-way analysis of variance with Tukey’s post hoc test. (H) Proposed model depicting ERFE’s mechanism of action: BMP2/6 heterodimeric ligand is secreted by liver endothelial cells and binds to the BMP receptor complex that has been proposed to contain two BMP type II receptors (RII), two type I receptors in the form of ALK3/ALK3 homodimers or ALK2/3 heterodimers, the coreceptor hemojuvelin (HJV), and possibly other interacting proteins, including HFE, transferrin receptor 2 (TFR2), and neogenin (Neo).10,14,17 Activated type I receptors phosphorylate SMAD1/5/8 proteins, which complex with SMAD4 and translocate to the nucleus to induce hepcidin transcription. In the context of erythropoietic drive, secreted ERFE binds to BMP2/6 to prevent BMP2/6 from binding and activating the BMP receptor complex, thereby suppressing hepcidin transcription.
To understand the in vivo observations, we took advantage of recent data that hemochromatosis severity was not exacerbated in double endothelial Bmp2 and Bmp6 knockout mice compared with single knockout mice,15 suggesting that BMP2 and BMP6 work collaboratively in vivo, potentially as a heterodimer. We thus examined whether heterodimeric BMP2/6 binds ERFE and its type I receptor utilization. We found that unlike BMP6, BMP2/6 preferentially used ALK3 for hepcidin induction (Figure 2D). Moreover, BMP2/6-mediated hepcidin induction was suppressed by ERFE (Figure 2E). As previously reported, 13 ERFE also inhibited hepcidin induction by BMP6. In contrast, although hepcidin levels were lower in cells treated with BMP2 and ERFE compared with BMP2 treatment alone, the fold-induction was not reduced after adjusting for lower basal hepcidin levels in ERFE-treated control mice (Figure 2E). This outcome suggests that ERFE does not functionally inhibit BMP2 at concentrations sufficient to inhibit BMP6 and BMP2/6. Finally, ERFE bound directly to BMP2/6 heterodimers and prevented BMP2/6 binding to ALK3 (Figure 2F-G; supplemental Figure 6). These data suggest that ERFE inhibits hepcidin by acting as a ligand trap for the physiological form of BMP ligand in vivo, BMP2/6, which preferentially utilizes ALK3 for signaling (Figure 2H). These results are consistent with the major role of ALK3 compared with ALK2 in hepcidin suppression by EPO/ERFE, and they help explain the more robust iron overload phenotype in Bmp6−/− and Alk3fl/fl;Alb-Cre mice compared with Alk2fl/fl;Alb-Cre mice.7,8,16
The online version of this article contains a data supplement.
Acknowledgments
The authors thank Tomas Ganz and Elizabeta Nemeth (UCLA) for graciously providing the HEK293T cells overexpressing ERFE.
This work was supported by National Institutes of Health, National Institute of Diabetes and Digestive and Kidney Diseases grant RO1-DK087727 and a Massachusetts General Hospital Research Scholar Award to J.L.B. A.U.S. was supported by grants from the German Research Foundation (Deutsche Forschungsgemeinschaft) STE 1895/4-2 and the Innovative Medical Research (Innovative Medizinische Forschung) (IMF 22 18 10) of the Medical Faculty, University of Muenster. C.-Y.W. is supported in part by a Cooley’s Anemia Foundation Research Fellowship, and L.T. is supported by the German Research Foundation (TR 1642/1-1).
Authorship
Contribution: C.-Y.W. designed and performed all cell culture experiments, interpreted data, created a graphical summary, and wrote the paper; Y.X. designed and performed all binding experiments, interpreted data, and edited the paper; L.T. and D.Y.D. performed mouse experiments, interpreted data, and edited the paper; X.X. assisted cell culture experiments and edited the paper; A.U.S. performed mouse experiments, oversaw the study, interpreted data, and wrote the paper; and J.L.B. conceived and oversaw the study, interpreted data, and wrote the paper.
Conflict-of-interest disclosure: J.L.B. has ownership interest in Ferrumax Pharmaceuticals and has received consulting fees from Keryx Biopharmaceuticals and Disc Medicine. The remaining authors declare no competing financial interests.
Correspondence: Jodie L. Babitt, Massachusetts General Hospital, 185 Cambridge St, CPZN-8208, Boston, MA 02114; e-mail: babitt.jodie@mgh.harvard.edu; or Andrea U. Steinbicker, University Hospital Muenster, Albert-Schweitzer Campus 1, Building A1, 48149 Muenster, Germany; e-mail: andrea.steinbicker@ukmuenster.de.