

Abstract
Iron-deficiency anemia (IDA) affects many infants in low- and middle-income countries (LMICs) and may impair cognitive development and adaptive immunity. Effective interventions to improve iron intakes for infants in LMICs are urgently needed. However, absorption of oral iron fortificants and supplements is low, usually <10%, and most of the iron passes into the colon unabsorbed. In randomized controlled trials, provision of iron to infants in LMICs adversely affects their gut microbiome and increases pathogenic Escherichia coli, gut inflammation, and diarrhea. To minimize these detrimental effects of iron, it is important to provide the lowest effective dosage and maximize fractional iron absorption. Prebiotic galacto-oligosaccharides and apo-lactoferrin may prove useful in iron formulations in LMICs because they increase absorption of fortificant iron and at the same time may mitigate the adverse effects of unabsorbed iron on the infant gut. Providing well-absorbed iron early in infancy may improve immune function. Recent data from a Kenyan birth cohort suggest IDA at the time of infant vaccination impairs the response to diphtheria, pertussis, and pneumococcus vaccines. A randomized trial follow-up study reported that providing iron to Kenyan infants at the time of measles vaccination increased antimeasles immunoglobulin G (IgG), seroconversion, and IgG avidity. Because IDA is so common among infants in LMICs and because the vaccine-preventable disease burden is so high, even if IDA only modestly reduces immunogenicity of vaccines, its prevention could have major benefits.
Learning Objectives
Understand the adverse effects of iron fortification on the infant gut microbiome, gut inflammation, and diarrhea and potential approaches to mitigate these adverse effects of iron
Understand the links between iron deficiency anemia, adaptive immunity, and infant vaccine response
Clinical case
A 10-week-old breastfeeding male infant presents to an outpatient clinic in southern rural Kenya for routine vaccination. The mother states that the infant, who was born with low birth weight, is irritable and not feeding well. The infant has had several bouts of fever and watery diarrhea over the past month. On clinical examination, he is afebrile, and chest auscultation is unremarkable. However, he is underweight, and his mucosae and conjunctiva are markedly pale. Malaria test is negative, but hemoglobin is 8.8 g/dL. What is the likely etiology for his anemia? What treatment will you provide? Is supplemental iron safe in this setting? Should iron-fortified foods be added to his diet? Will supplemental iron aggravate the infant’s diarrheal episodes?
Introduction
Iron-deficiency anemia (IDA) affects >1 billion people worldwide and is one of the five leading global causes of years lived with disability.1 IDA is particularly prevalent in low- and middle-income countries (LMICs), and in Africa, 62% of preschool children are anemic,2 mainly because of iron deficiency. Infants in LMICs have particularly high rates of IDA during the weaning period because high iron needs for growth and erythropoiesis typically are not covered by the low amounts of iron present in breastmilk and plant-based complementary foods.3 Defining iron status in very young infants is challenging because of rapid changes in iron metabolism during the first few months after birth.4 IDA in infants and young children may impair cognitive development, and this impairment may be irreversible or only partially reversible by iron repletion.3 Because IDA during infancy is common and can lower the intelligence quotient, it has serious health and economic costs and may hinder national development.5 Moreover, recent findings have linked IDA to impairments in adaptive immunity and, possibly, vaccine response.6 Thus, effective, sustainable, and safe interventions to improve iron intakes in infants in LMICs are urgently needed.3
Iron fortification, the gut microbiome, and diarrhea
Dietary iron absorption is tightly regulated in humans because there is no active pathway for iron excretion.7 Iron absorption from iron-fortified foods or iron supplements is generally low, typically <10%.3 In LMICs, iron absorption is likely to be even lower, because common infections and inflammation increase plasma hepcidin, which reduces iron absorption.7 Thus, the majority of oral iron passes unabsorbed into the colon, where the iron is available to gut microbes. Iron plays a key role in replication and virulence (eg, adhesion, invasion, and induction of virulence factors) of many enteric gram-negative bacteria (eg, Salmonella, pathogenic Escherichia coli).8,9 In contrast, beneficial commensal gut bacteria, such as Bifidobacteriaceae and Lactobacillaceae, which provide an important barrier against colonization by enteropathogens, need very little or no iron.10 Thus, an increase in unabsorbed dietary iron entering the colon may tip the balance toward growth of potential enteropathogens over important commensal “barrier” strains. Systemically, hepcidin-mediated iron sequestration withholds iron from invading microorganisms and plays a key role in innate immune responses to infection.11 Similarly, hepcidin production by mucosal dendritic cells, independent of systemic iron or hepcidin levels, may sequester iron into colonic myeloid cells; failure to sequester iron locally results in dysbiosis, increases microbial translocation, and worsens gut inflammation.12
The World Health Organization recommends providing micronutrient powders (MNPs) containing 12.5 mg of iron to infants aged 6 to 23 months in areas where the anemia prevalence is high.13 However, these iron-containing MNPs are not entirely safe for infants,14 because they increase diarrhea risk. A systematic review reported a 15% increased risk for diarrhea (risk ratio 1.15; 95% confidence interval, 1.06-1.26) with iron dosages given at ≥80% of the recommended daily intake.15 A review of potential mechanisms linking oral iron dosages to diarrhea suggested an increase in Enterobacteriaceae and pathogenic E. coli or a decrease in Bifidobacteriaceae may play a role.16 Pathogenic E. coli is an important cause of diarrhea17 and bacteremia18 among African infants. Diarrhea is a leading cause of child mortality in LMICs; the World Health Organization estimates that diarrhea contributes to 19% of under-5 deaths globally.19
Recent randomized controlled trials assessed the impact of iron-containing MNPs on the infant gut microbiota, gut inflammation, and diarrhea in Kenyan infants.20,21 Infants in the first trial (n = 115) consumed MNPs containing 2.5 or 12.5 mg iron or without iron daily for 4 months.20 Iron deficiency was defined as a low serum ferritin (SF) or an elevated soluble transferrin receptor (sTfR) or erythrocyte zinc protoporphyrin; screening for hemoglobinopathies was not performed. There was a significant positive treatment effect of the MNP containing 12.5 mg iron on SF, sTfR, and zinc protoporphyrin (for all, P < .05) but no significant effect of the MNP containing 2.5 mg iron. At baseline, the gut microbiota of the infants harbored a high abundance of beneficial Bifidobacteriaceae (63%). At the same time, many were carrying potential pathogens: 65% were positive for enteropathogenic E. coli, 49% for enterotoxigenic E. coli producing heat-labile toxin, 57% for Clostridium difficile, and 22% for Salmonella. In the iron groups compared with the no-iron control groups, there was a significant increase in Enterobacteriaceae, particularly Escherichia/Shigella spp., the enterobacteria/bifidobacteria ratio, and Clostridium (for all, P < .05). Moreover, there were higher abundances of pathogenic E. coli at endpoint in the iron groups (6.0 ± 0.5 log gene copy number/g feces) compared with the no-iron groups (4.5 ± 0.5) (P = .029) (Figure 1). In addition, fecal calprotectin (a marker of gut inflammation) was significantly higher in the iron compared with the no-iron control group (Figure 2), and diarrheal rates were higher.20 This study was the first to demonstrate that provision of iron to African infants adversely affects gut microbiome composition and increases gut inflammation.
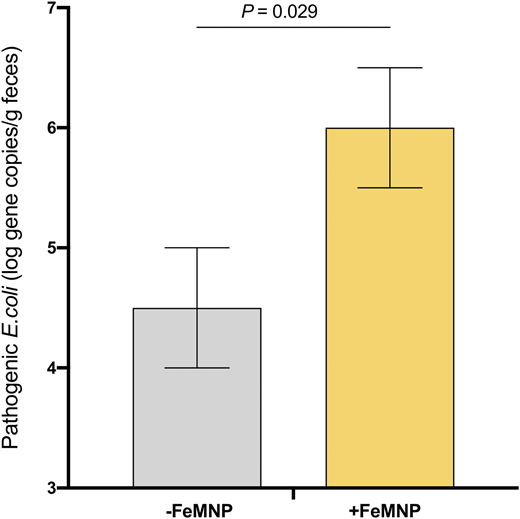
Abundance of pathogenic E. coli at endpoint, by group, in Kenyan infants (n = 115) receiving daily for 4 months an MNP containing either no iron (−FeMNP) or 2.5 mg or 12.5 mg iron (+FeMNP). Univariate general linear models with baseline values as covariates were used to estimate the intervention effect. Adapted from Jaeggi et al.20
Abundance of pathogenic E. coli at endpoint, by group, in Kenyan infants (n = 115) receiving daily for 4 months an MNP containing either no iron (−FeMNP) or 2.5 mg or 12.5 mg iron (+FeMNP). Univariate general linear models with baseline values as covariates were used to estimate the intervention effect. Adapted from Jaeggi et al.20
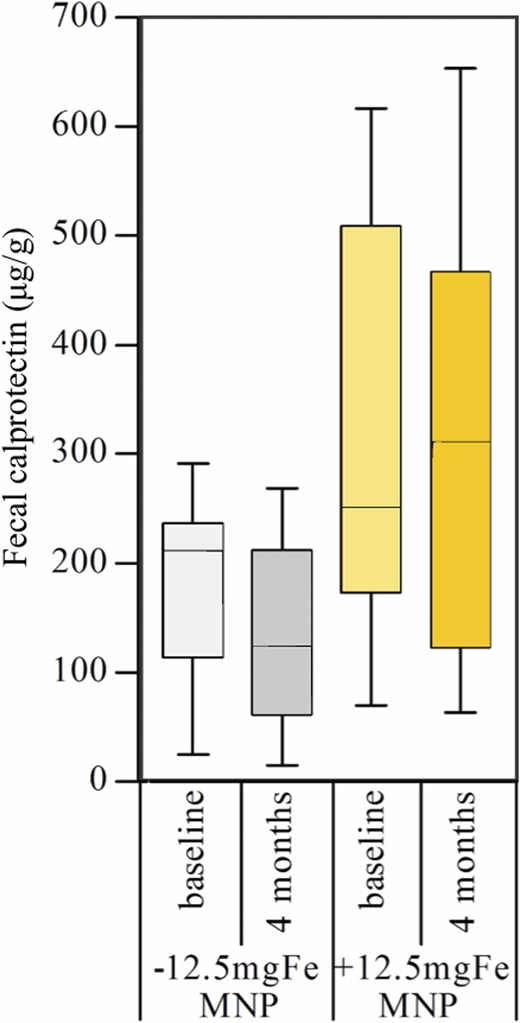
Fecal calprotectin levels at baseline and 4 months, by group, in Kenyan infants (n = 115) receiving daily for 4 months an MNP containing no iron (−12.5mgFeMNP) or containing 12.5 mg iron (+12.5mgFeMNP). At endpoint, fecal calprotectin was significantly higher in the +12.5mgFeMNP group than in the −12.5mgFeMNP group (P = .008). Differences were investigated via general linear models with baseline variables as covariates. Boxplots are shown with the 10th to 90th percentiles. Adapted from Jaeggi et al.20
Fecal calprotectin levels at baseline and 4 months, by group, in Kenyan infants (n = 115) receiving daily for 4 months an MNP containing no iron (−12.5mgFeMNP) or containing 12.5 mg iron (+12.5mgFeMNP). At endpoint, fecal calprotectin was significantly higher in the +12.5mgFeMNP group than in the −12.5mgFeMNP group (P = .008). Differences were investigated via general linear models with baseline variables as covariates. Boxplots are shown with the 10th to 90th percentiles. Adapted from Jaeggi et al.20
In a subsequent 4-month randomized controlled trial in the same study area,21 infants aged 6.5 to 9.5 months (n = 155) were randomly assigned to receive daily an MNP without iron (control group), the identical MNP but with 5 mg iron (Fe group), or the identical MNP as the Fe group but with 7.5 g galacto-oligosaccharides (GOS) (FeGOS group). Iron deficiency was defined as a low SF or an elevated sTfR; screening for hemoglobinopathies was not performed. At 4 months, compared with the control group, there was significant improvement in PF and sTfR in the Fe and FeGOS groups (P < .001 for both). GOS is a nondigestible prebiotic carbohydrate that enters the colon intact and selectively enhances the growth of beneficial commensal Bifidobacterium and Lactobacillus.22 Prebiotics may protect from colonization and overgrowth of potential enteric pathogens by increasing colonization resistance, increasing production of short chain fatty acids, and decreasing colonic luminal pH.22 The addition of GOS to the iron-containing MNP mitigated most of the adverse effects of iron on the infant gut microbiota.21 After 4 months of intervention, compared with the Fe group, in the FeGOS group there was a higher abundance of Bifidobacterium and Lactobacillus and lower abundance of Clostridiales (Figure 3). Remarkably, there were no significant differences in the abundances of Bifidobacterium, Lactobacillus, Enterobacteriaceae, Clostridiales, or Bacteroidetes between the control and FeGOS groups (Figure 3). After 3 weeks of intervention there were lower abundances of the sum of virulence and toxin genes of all pathogens in the FeGOS group compared with both the control group and the Fe group (Figure 4). Also, there were significantly lower abundances of the sum of virulence and toxin genes of pathogenic E. coli in the FeGOS group compared with the control group. At 4 months, plasma intestinal fatty acid–binding protein (a biomarker of enterocyte damage) was significantly higher in the Fe group than in the control group but was not higher in the FeGOS group when compared with the control group.21 Results of a recent study of Swedish infants also suggest that iron-fortified formula and iron supplements may adversely affect the gut microbiome and that prebiotic GOS in infant formula may be protective.23
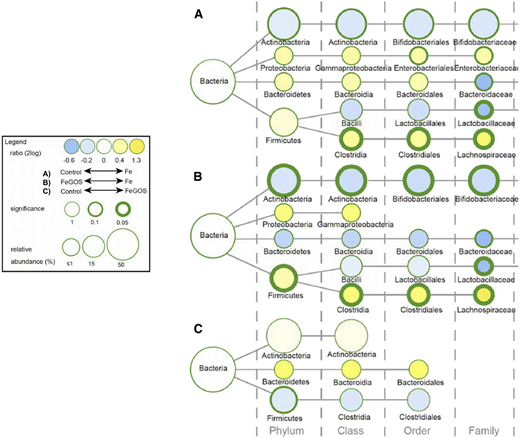
Differences in the gut microbiota composition among Kenyan infants (n = 155) after receiving daily for 4 months an MNP without iron (control group), with 5 mg of iron (Fe group), or with 5 mg iron and 7.5 g galacto-oligosaccharides (FeGOS group). Nodes represent taxa; edges link the different taxonomic levels. Node sizes correspond to the relative taxa abundance (%). The fold difference is calculated as the 2log of the ratio of the relative abundance between groups. Mann–Whitney U tests were used for statistical comparisons. Adapted from Paganini et al.21
Differences in the gut microbiota composition among Kenyan infants (n = 155) after receiving daily for 4 months an MNP without iron (control group), with 5 mg of iron (Fe group), or with 5 mg iron and 7.5 g galacto-oligosaccharides (FeGOS group). Nodes represent taxa; edges link the different taxonomic levels. Node sizes correspond to the relative taxa abundance (%). The fold difference is calculated as the 2log of the ratio of the relative abundance between groups. Mann–Whitney U tests were used for statistical comparisons. Adapted from Paganini et al.21
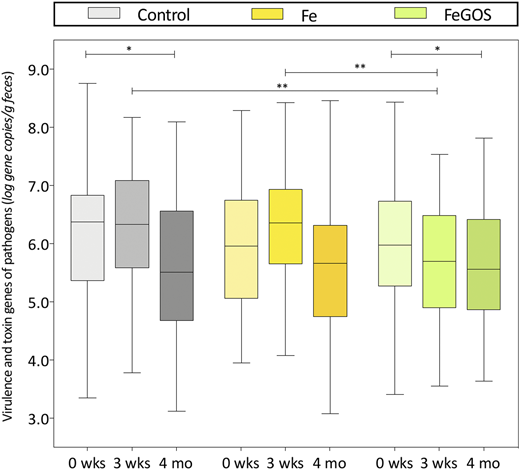
Abundances of the sum of virulence and toxin genes of 10 pathogens at baseline (0 weeks), 3 weeks, and 4 months, by group in Kenyan infants (n = 155) consuming daily an MNP containing either no iron (control), 5 mg of iron (Fe), or 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS). Significance is expressed as the P value of a Wilcoxon rank-sum test, *P < .05, **P < .01. Boxes show the median and 25th and 75th percentiles; whiskers show the range. Adapted from Paganini et al.21
Abundances of the sum of virulence and toxin genes of 10 pathogens at baseline (0 weeks), 3 weeks, and 4 months, by group in Kenyan infants (n = 155) consuming daily an MNP containing either no iron (control), 5 mg of iron (Fe), or 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS). Significance is expressed as the P value of a Wilcoxon rank-sum test, *P < .05, **P < .01. Boxes show the median and 25th and 75th percentiles; whiskers show the range. Adapted from Paganini et al.21
Maximizing iron absorption from iron supplements and fortificants in infancy
Because of the detrimental effects of unabsorbed iron on the gut microbiome of African infants,19 it is important to provide the lowest effective dosage by maximizing fractional iron absorption. Recent data demonstrate that prebiotic GOS may increase iron absorption when added to iron-containing MNPs.16 Kenyan infants (n = 50; aged 6–14 months) consumed maize porridge that was fortified with an MNP containing iron (5 mg) and GOS (7.5 g) or the same MNP without GOS each day for 3 weeks. Then all infants were provided an isotopically labeled maize porridge and MNP test meal containing iron either as a mixture of ferrous fumarate and sodium iron ethylenediaminetetraacetate or as ferrous sulfate. Iron absorption was measured as the erythrocyte incorporation of stable isotopes. GOS consumption by the infants significantly increased iron absorption by +62% from the MNP containing ferrous fumarate and sodium iron ethylenediaminetetraacetate.16
Lactoferrin is a glycosylated protein highly concentrated in human milk and can exist in an apo (iron-free) state or can bind two ferric ions with very high affinity, forming holo-lactoferrin.24 Because lactoferrin binds iron with high affinity at the pH of the infant small and large intestine, it may be a bacteriostatic agent for withholding iron from enteropathogens, whose growth and virulence depend on an adequate iron supply.24 Whether lactoferrin also binds iron to facilitate its absorption remains uncertain, but a lactoferrin receptor has been identified on enterocytes, which may mediate iron transport from lactoferrin into mucosal cells.25 In a recent randomized crossover trial, Kenyan infants (n = 25, mean age 4 months) were fed multiple stable iron isotopes in ferrous sulfate–fortified test meals with and without bovine apo-lactoferrin, with iron absorption quantified by measurement of erythrocyte iron incorporation.26 The addition of apo-lactoferrin significantly increased iron absorption by +56% (Figure 5). Iron absorption was also measured from intrinsically labeled holo-lactoferrin, and it was comparable to iron absorption from ferrous sulfate alone (Figure 5). Intervention studies of infants assessing the effect of supplemental lactoferrin on iron status are equivocal, but the use of lactoferrin in iron-fortified infant formula is associated with a reduced incidence and duration of diarrhea.27 Supplemental lactoferrin has also demonstrated benefits on iron status in pregnant women.28 Therefore, prebiotic GOS and bovine apo-lactoferrin may prove useful in iron formulations for infants in LMICs because they both increase absorption of iron and potentially mitigate the adverse effects of unabsorbed iron on the gut microbiome.
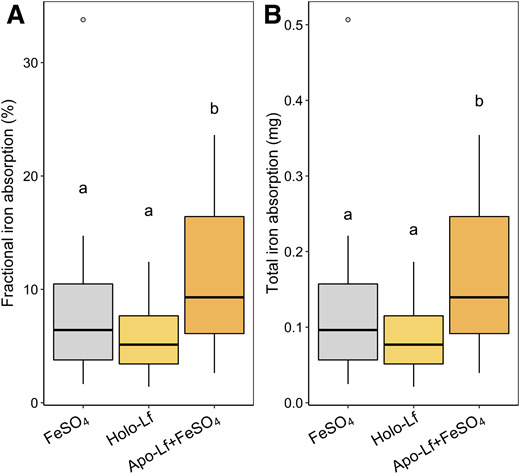
Fractional iron absorption (%) (A) and total iron absorption (mg) (B) in Kenyan infants (n = 25) from maize porridge containing a mixture of 1.5 mg iron as54Fe-labeled FeSO4; as58Fe-labeled FeSO4+ 1.41 g apo-lactoferrin; and as intrinsically57Fe-labeled holo-lactoferrin containing 1.41 g lactoferrin. The horizontal lines show the geometric means, and the whiskers extend from –SD to +SD. Data analyzed with repeated-measures analysis of covariance with Bonferroni post hoc analysis. Geometric means without a common letter differ, P < .05. FeSO4, ferrous sulfate; Lf, lactoferrin. Adapted from Mikulic et al.26
Fractional iron absorption (%) (A) and total iron absorption (mg) (B) in Kenyan infants (n = 25) from maize porridge containing a mixture of 1.5 mg iron as54Fe-labeled FeSO4; as58Fe-labeled FeSO4+ 1.41 g apo-lactoferrin; and as intrinsically57Fe-labeled holo-lactoferrin containing 1.41 g lactoferrin. The horizontal lines show the geometric means, and the whiskers extend from –SD to +SD. Data analyzed with repeated-measures analysis of covariance with Bonferroni post hoc analysis. Geometric means without a common letter differ, P < .05. FeSO4, ferrous sulfate; Lf, lactoferrin. Adapted from Mikulic et al.26
Iron deficiency anemia, adaptive immunity, and infant vaccine response
Immunization programs in LMICs have achieved high coverage, yet 1 in 5 children worldwide are not fully protected, contributing to 1.5 million child deaths yearly from vaccine-preventable diseases.29 Vaccines often underperform in LMICs.30 For example, effectiveness of measles vaccine is generally <75% in Sub-Saharan Africa.31 Why vaccines underperform in LMICs remains uncertain,30 but new data suggest iron deficiency (ID) may play a role.32,33
Previous reviews34 have suggested multiple mechanisms by which iron status might influence adaptive immunity. ID in mice attenuates T-cell–dependent and T-cell–independent antigen-specific antibody responses and impairs cyclin E1 induction and S-phase entry during B-cell proliferation.32 In some animal and cell models, ID reduces the proportion of mature T cells and impairs T-cell activation and proliferation.34 In humans, studies on ID and immune function show varying results depending on what aspect of immunity is measured and the severity of ID, age, and underlying nutritional status.34 Iron uptake via transferrin receptor 1 is essential for lymphocyte development, and clinically, a homozygous mutation in transferrin receptor 1 causes severe immunodeficiency in children and reduced numbers of circulating memory B cells.33 In vitro, this mutation prevented T- and B-cell proliferation, and addition of iron citrate in vitro rescued the proliferative defect.33 Thus, adequate iron availability may be critical for adaptive immunity. If ID during infancy limits iron availability to responding lymphocytes,32 this could impair response to vaccination. Recent studies showing impaired antigen-specific immune responses in hypoferremic mice support this concept (Joe Frost et al, University of Oxford, written personal communication, June 25, 2020).
IDA is particularly common among infants age <1 year in Sub-Saharan Africa: In southern Kenya, 70% to 75% of infants are anemic at the time they receive their routine vaccinations.35 Term infants born to iron-sufficient mothers should have adequate birth iron stores to cover iron requirements for the first 4 to 6 months.36 However, in Sub-Saharan Africa, 46% of pregnant women have IDA,37 reducing maternal–fetal iron transfer.36 The umbilical cord is often clamped too early, and 15% to 25% of newborns have low birthweight.38 These factors sharply reduce newborn iron stores: It is estimated that body iron is 40% to 50% lower in newborns who are low birthweight or whose mothers were anemic during pregnancy.36 Low iron stores at birth, together with frequent infections increasing serum hepcidin35 and diarrhea or intestinal parasites causing blood loss, result in many infants depleting their iron stores within 3 to 4 months after birth.
Two recent studies in southern coastal Kenya suggest that IDA during infancy impairs vaccine response.6 A birth cohort study assessed whether anemia or ID at time of vaccination predicted vaccine response to 3-valent oral polio, diphtheria–tetanus–whole cell pertussis–Haemophilus influenzae type b vaccine, 10-valent pneumococcal conjugate vaccine, and measles vaccine. Primary outcomes were anti-vaccine immunoglobulin G (IgG) and seroconversion at age 24 weeks and 18 months. A total of 573 infants were enrolled at birth, and 303 completed the study.6 More than half of infants were already anemic at age 10 weeks. With sex, birthweight, anthropometric indices (z scores for length and weight), and maternal antibodies controlled for, hemoglobin at time of vaccination was the strongest positive predictor of antidiphtheria and anti-pertussis-IgG at 24 weeks and 18 months, antipertussis filamentous hemagglutinin-IgG at 24 weeks, and anti–pneumococcus 19 IgG at 18 months (for all, P < .05). Anemia and serum transferrin receptor at time of vaccination were the strongest predictors of seroconversion against diphtheria and pneumococcus 19 at 18 months (for both, P < .05). In a randomized trial cohort follow-up,6 infants (n = 155) received an MNP with 5 mg iron daily or an MNP without iron for 4 months starting at age 7.5 months and received measles vaccine at age 9 months. Vaccine response was measured at age 11.5 months and 4.5 years. Compared with infants who did not receive iron, those who received iron at time of vaccination had higher antimeasles IgG, seroconversion, and IgG avidity (for all, P < .05) at age 11.5 months (Figure 6).6
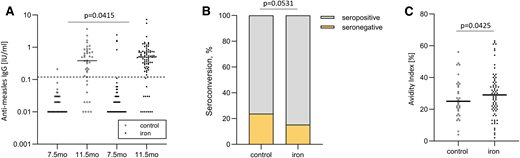
In a randomized controlled trial follow-up study of Kenyan infants (n = 155), antimeasles serum IgG concentrations, seroconversion, and IgG avidity in the control and iron groups. The iron group received 5 mg iron daily for 4 months at time of measles vaccination. (A) Anti-measles serum IgG concentrations at age 7.5 and 11.5 months (at baseline and end of intervention), (B) seroconversion at age 11.5 months (at end of intervention), and (C) IgG avidity at age 11.5 months (at end of intervention). Data analyzed with random intercept linear mixed effect models with Bonferroni corrected multiple comparisons. Adapted from Stoffel et al.6
In a randomized controlled trial follow-up study of Kenyan infants (n = 155), antimeasles serum IgG concentrations, seroconversion, and IgG avidity in the control and iron groups. The iron group received 5 mg iron daily for 4 months at time of measles vaccination. (A) Anti-measles serum IgG concentrations at age 7.5 and 11.5 months (at baseline and end of intervention), (B) seroconversion at age 11.5 months (at end of intervention), and (C) IgG avidity at age 11.5 months (at end of intervention). Data analyzed with random intercept linear mixed effect models with Bonferroni corrected multiple comparisons. Adapted from Stoffel et al.6
These are the first prospective data from Africa assessing the impact of anemia and ID at the time of vaccination on response to a range of pediatric vaccines. These data suggest that anemia or ID at the time of infant vaccination may impair the response to diphtheria, pertussis, and pneumococcus vaccines and that improving iron status may improve response to measles vaccine.6 These findings must be confirmed in other prospective cohorts and larger intervention trials. Powerful emerging techniques combining mass cytometry and systems-level omics tools39 may allow identification of the mechanisms underlying the effects of iron status on the infant immune system and response to vaccines. If confirmed, these findings argue strongly for early detection of IDA in infancy and its correction with effective iron treatment, as well as ensuring adequate iron status in pregnant women. Because anemia is so common in African infants and because the vaccine-preventable disease burden is so high,29 even if IDA only modestly reduces the immunogenicity of childhood vaccines, its prevention could have major benefits.
Conclusions
Returning to our clinical case, the likely etiology of anemia in this infant is iron deficiency due to several factors: low birthweight, resulting in low birth iron stores; exclusive breastfeeding, providing only very low levels of dietary iron; frequent diarrhea, resulting in increased gastrointestinal iron losses; and common infections, increasing serum hepcidin, which reduces iron absorption. According to protocols from the Kenyan Ministry of Health,40 the infant should be treated with oral iron syrup at a dosage of 2 to 6 mg/kg body weight for 30 days, and exclusive breastfeeding should continue until age 6 months. Provided at time of vaccination, this additional iron may improve the infant’s vaccine response. However, the mother should be told that the iron will probably darken the infant’s stool, and it may cause diarrhea. If the latter occurs, the iron syrup should be discontinued and the infant be brought back to clinic for treatment. At age 6 months, if anemia persists, complementary foods rich in iron, such as eggs and green leafy vegetables, should be regularly fed to the infant. In addition, iron-fortified MNPs can be given to the infant; if possible, these should be given with a prebiotic to reduce adverse effects on the gut microbiome and risk for diarrhea.
Correspondence
Laboratory of Human Nutrition, ETH Zurich, Swiss Federal Institute of Technology, LFVD20, Schmelzbergstrasse 7, CH-8092 Zurich, Switzerland, e-mail: michael.zimmermann@hest.ethz.ch
