

Key Points
HLA-DRB3*01:01 has a dose-dependent impact on HPA-1a antibody levels in HPA-1a–immunized women giving birth to an HPA-1a–positive child.
HLA-DRB3*01:01 has a dose-dependent impact on the neonatal platelet counts in HPA-1a–positive children born of HPA-1a–immunized women.

Abstract
HLA-DRB3*01:01 is a predisposing factor for human platelet antigen 1a (HPA-1a) immunization, which is responsible for most cases of fetal and neonatal alloimmune thrombocytopenia. The aim of this study was to investigate if the HLA-DRB3*01:01 allele imposes a dose-dependent effect on anti-HPA-1a levels and neonatal platelet counts. One hundred and thirty HPA-1a–immunized women were divided into 3 groups: HLA-DRB3*01:01 negative, HLA-DRB3*01:01 hemizygous or heterozygous, and HLA-DRB3*01:01 homozygous. The dose of the HLA-DRB3*01:01 allele was determined by sequencing exon 2 of the HLA-DRB3 gene followed by HLA-DRB3 and HLA-DRB1 typing of selected samples. Anti-HPA-1a levels at time of delivery and neonatal platelet counts were compared among groups. There was a significant dose-dependent effect of the HLA-DRB3*01:01 allele on anti-HPA-1a levels (global P value [Pglobal] = .0032). Median (range) anti-HPA-1a levels were 1.5 IU/mL (0.0-19.0 IU/mL), 21.1 IU/mL (0.0-1967 IU/mL), and 43.7 IU/mL (1.0-980 IU/mL) in women with 0, 1, and 2 copies of the HLA-DRB3*01:01 allele, respectively. There was also a significant, but opposite, dose-dependent effect of the mother’s HLA-DRB3*01:01 allele on the platelet count of the newborn (Pglobal = .0155). Median (range) neonatal platelet counts were 241 × 109/L (59 × 109/L to 393 × 109/L), 107 × 109/L (4 × 109/L to 387 × 109/L) and 32 × 109/L (4 × 109/L to 352 × 109/L) for newborns of mothers with 0, 1, and 2 copies of the HLA-DRB3*01:01 allele, respectively. Thus, the HLA-DRB3*01:01 allele exhibits a dose-dependent impact on maternal anti-HPA-1a levels in HPA-1a–immunized women.
Introduction
Fetal and neonatal alloimmune thrombocytopenia (FNAIT) is the platelet equivalent to hemolytic disease of the fetus and newborn. Maternal immunoglobulin G (IgG) alloantibodies against paternally inherited antigens can traverse the placenta and sensitize fetal platelets, which are then phagocytosed by splenic macrophages in the fetal spleen. FNAIT is a rare disease that affects ∼1 in 1000 fetuses/newborns.1 Most cases are benign and result in temporary thrombocytopenia with or without clinical symptoms of bleeding. One of the most feared consequences of FNAIT is intracranial hemorrhage (ICH), which may lead to intrauterine death or lifelong disability, which have been estimated to occur in ∼1 of 10 000 newborns.2 Given an annual birth rate of ∼10 million in Europe and North America, this incidence rate translates to ∼1000 yearly cases of FNAIT-associated ICH.
The most common and severe cases of FNAIT in whites are caused by antibodies against the human platelet antigen 1a (HPA-1a). The epitope is located on the β3 integrin (GPIIIa),3 which is present on the platelet surface from gestational week 16.4 HPA-1a and HPA-1b are determined by 2 allelic variants of the β3 integrin molecule, which results in a substitution of proline for leucine at amino acid residue 33 of the β3 integrin molecule.5
For >2 decades, it has been known that the majority of HPA-1a–negative women, who develop HPA-1a antibodies during or after a pregnancy with an HPA-1a–positive child, are HLA-DRB3*01:01 positive.6-12 As the HLA-DRB3 gene is only present in certain HLA-haplotypes, an individual can either lack HLA-DRB3 or have 1 or 2 expressed copies of this gene. There are currently (October 2018) 175 HLA-DRB3 alleles,13 of which HLA-DRB3*01:01 is one of the most common (ie, 27.6% of whites are positive for this HLA-DRB3 allele).14 The risk of HPA-1a immunization after delivery of an HPA-1a–positive child is 12.7% in women who are HLA-DRB3*01:01 positive but only 0.5% in women who lack this HLA allele.15 As expression of HLA-DRB3*01:01 is codominant, it would be interesting to know if the HLA-DRB3*01:01 allele imposes a dose-dependent effect on the level of potentially pathogenic maternal anti-HPA-1a antibodies and/or influences the neonatal platelet count. The current study is the first to address this question in a cohort of women prospectively identified as HPA-1a immunized.
Methods
Patients who previously were included in the Norwegian screening and intervention study to reduce morbidity and mortality caused by anti-HPA-1a–associated FNAIT7,16 were invited to take part in the present study. More than 100 000 pregnant women were HPA-1a typed in the previous screening study. Those who were HPA-1a negative were offered screening for HPA-1a antibodies approximately every month during pregnancy. Women who were found to be HPA-1a immunized were offered clinical follow-up at regular intervals at a center for maternal-fetal medicine and delivery by Caesarean section 2 to 4 weeks prior to term. HPA-1a–negative platelets were ready for transfusion at the operation theater, in case the newborn had clinical signs of bleeding or were thrombocytopenic.
Of the women who participated in the Norwegian screening and intervention study, 130 consented to participate in the current study, which entailed extended HLA typing of the women who previously took part in Norwegian FNAIT study. The study was conducted according to the principles of the Declaration of Helsinki and was approved by the Regional Committee for Medical and Health Research Ethics in North of Norway (consent 05/01500, 11/5-2006).
Of the 130 women included in the present study, 15 had been included twice in the Norwegian screening and intervention study.7,16 Two of these 15 women gave birth to 1 HPA-1a–positive child and 1 HPA-1a–negative child. In this study, we only included the mother-child pairs in which the children were HPA-1a positive. The other 13 women had 2 HPA-1a–positive children each, and for these cases, we only included the first mother-child pair in this study. None of the HPA-1a–immunized women were treated with high-dose IV IgG during pregnancy. Data regarding the women’s obstetric history and time of delivery, as well as the sex distribution and birth weight of the children, are given in Tables 1 and 2.
Number of women in the 3 categories with different numbers of previous pregnancies, and number of women with a previous history of FNAIT
| Dose of HLA-DRB3*01:01 | No. of previous pregnancies | Previous history of FNAIT | ||||
|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | ||
| No. of DRB3*01:01-negative women | 0 | 1 | 9 | 1 | 0 | 0 |
| No. of DRB3*01:01 hemizygous or heterozygous women | 10 | 44 | 32 | 14 | 6 | 8 |
| No. of DRB3*01:01 homozygous women | 2 | 4 | 2 | 3 | 2 | 0 |
| Dose of HLA-DRB3*01:01 | No. of previous pregnancies | Previous history of FNAIT | ||||
|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | ||
| No. of DRB3*01:01-negative women | 0 | 1 | 9 | 1 | 0 | 0 |
| No. of DRB3*01:01 hemizygous or heterozygous women | 10 | 44 | 32 | 14 | 6 | 8 |
| No. of DRB3*01:01 homozygous women | 2 | 4 | 2 | 3 | 2 | 0 |
Time of delivery as well as sex distribution and birth weight of the children born of the women in the 3 categories
| Dose of HLA-DRB3*01:01 | Time of delivery (no. of days before term), mean (SD) | Birth weight, mean (SD), g | Sex distribution (no. of girls/no. of boys) |
|---|---|---|---|
| DRB3*01:01-negative women | 20.9 (9.1) | 3250 (489) | 4/7 |
| DRB3*01:01 hemizygous or heterozygous women | 22.7 (9.5) | 2979 (506)* | 60/48* |
| DRB3*01:01 homozygous women | 23.3 (4.8) | 2907 (363) | 8/5 |
| Dose of HLA-DRB3*01:01 | Time of delivery (no. of days before term), mean (SD) | Birth weight, mean (SD), g | Sex distribution (no. of girls/no. of boys) |
|---|---|---|---|
| DRB3*01:01-negative women | 20.9 (9.1) | 3250 (489) | 4/7 |
| DRB3*01:01 hemizygous or heterozygous women | 22.7 (9.5) | 2979 (506)* | 60/48* |
| DRB3*01:01 homozygous women | 23.3 (4.8) | 2907 (363) | 8/5 |
These data include 2 pairs of twins.
Platelet count was assessed in all the newborns immediately after delivery, and this value was used for the statistical analyses. Two women gave birth to twins with platelet counts of 237/295 and 34/33, respectively. These cases were each considered as 1 mother-child pair, and the mean platelet counts of 266 and 34 were used for calculating the statistics.
A sample for determination of maternal anti-HPA-1a level was collected in relation to delivery. An in-house quantitative monoclonal antibody immobilization of platelet antigen (MAIPA) assay was used for quantification of anti-HPA-1a as previously described.17-19 Briefly, a sample from 1 woman with a high level of anti-HPA-1a antibodies (a secondary standard of 15 IU/mL when compared with the international anti-HPA-1a standard, NIBSC product code 03/152, Hertfordshire, United Kingdom) was used as a reference. This reference sample was diluted twofold from 1:2 to 1:64 to create a linear standard curve. The MAIPA assay was carefully validated and found to be robust, as the validation samples were quantified with acceptable accuracy and precision. In addition, interlaboratory reproducibility has been shown to be satisfactory.18 The same MAIPA protocol was used for quantification of all included samples. Samples from women with high levels of antibodies were further sequentially diluted in twofold steps in phosphate-buffered saline with 2% bovine serum albumin until the measurement was within the linear part of the standard curve. A negative sample was also run in parallel.
HLA-DRB3 typing was performed by sequencing exon 2 of the HLA-DRB3 gene as the first step to determine the dose of the HLA-DRB3*01:01 allele. The intron-located primers previously described by Kotsch et al20 were used for polymerase chain reaction (PCR). The PCR products were sequenced in an ABI PRISM 3730 DNA analyzer (Applied Biosystems, Foster City, CA). DNA from women whose sequencing results were difficult to interpret and HLA-DRB3–negative women were retyped using HLA-DRB3 PCR-sequence–specific primers (GenoVision, Vienna, Austria).
Due to linkage disequilibrium, the HLA-DRB1 allele groups *03, *11, *12, *13, and *14 are all strongly associated with HLA-DRB3, while the rest of the HLA-DRB1 allele groups almost never coexist with the HLA-DRB3 alleles. For this reason, HLA-DRB1 typing was performed on cases where sequencing alone could not resolve the dose of HLA-DRB3*01:01 (eg, individuals who were positive for DRB3*01:01, but not other DRB3 alleles). The DRB1 typing was first done by first-field resolution HLA typing and subsequently by second-field resolution HLA typing for selected samples using PCR-sequence–specific primers (GenoVision, Vienna, Austria).
Since maternal anti-HPA-1a levels and neonatal platelet counts were not normally distributed, the Kruskal-Wallis test was used for comparisons among the 3 groups of mother-child pairs (ie, women who were negative, hemizygous or heterozygous, and homozygous for the HLA-DRB3*01:01 allele). For this global test, a 2-tailed P < .05 was considered significant. The Mann-Whitney U test was used for comparisons between 2 groups of mother-child pairs. Since post-hoc testing implied 2 comparisons, 1-tailed P < .025 was considered significant. As the global test showed a clear dose effect, it was considered justified to apply 1-tailed tests for the post-hoc analyses. Since time of delivery (measured in days before term) and birth weight were normally distributed, mean and standard deviation (SD) were used for descriptive statistics. The statistical calculations were performed using MedCalc Statistical Software version 18.11 (MedCalc Software bvba, Ostend, Belgium; https://www.medcalc.org; 2018).
Results
The number of patients who were tested by the different testing strategies is given in Table 3. Sequencing of exon 2 of the HLA-DRB3 gene was performed in 128 women. Seven women apparently did not have an HLA-DRB3 gene. HLA-DRB3 typing of these samples confirmed absence of the HLA-DRB3 gene. Two women were HLA-DRB3*01:01 negative but expressed HLA-DRB3*02:02. In 19 women, HLA-DRB3*01:01 coexisted with another HLA-DRB3 allele. Interpretation of these samples was straightforward: 9 women were HLA-DRB3*01:01 negative, and 19 were heterozygous for the HLA-DRB3*01:01 allele.
Number of women analyzed by each of the 7 different typing strategies (rows) and final conclusion regarding their dose of HLA-DRB3*01:01
| Sequencing exon 2 of the HLA-DRB3 gene | HLA-DRB3 typing* | First-field resolution HLA-DRB1 typing* | Second-field resolution HLA-DRB1 typing* | Conclusion | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRB3*01:01 negative | One DRB3 allele | DRB3*01:01 in combination with another DRB3 allele | DRB3 negative | DRB3 allele other than *01:01 | No alleles associated with DRB3 | Only 1 allele associated with DRB3 | Two alleles associated with DRB3 | Two alleles associated with DRB3*01:01 | DRB3*01:01 negative | One dose of DRB3*01:01 | Two doses of DRB3*01:01 | |
| Only DRB3*01:01 | DRB3 allele other than *01:01 | |||||||||||
| 7 | — | — | — | 7 | — | — | — | — | — | 7 | — | — |
| — | — | 2 | — | — | — | — | — | — | — | 2 | — | — |
| — | — | — | 19 | — | — | — | — | — | — | — | 19 | — |
| — | 87 | — | — | — | — | — | 87 | — | — | — | 87 | — |
| — | 13 | — | — | — | — | — | — | 13 | 13 | — | — | 13 |
| — | — | — | — | — | — | 1 | — | — | — | 1 | — | — |
| — | — | — | — | — | 1 | — | — | — | — | 1 | — | — |
| Sequencing exon 2 of the HLA-DRB3 gene | HLA-DRB3 typing* | First-field resolution HLA-DRB1 typing* | Second-field resolution HLA-DRB1 typing* | Conclusion | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRB3*01:01 negative | One DRB3 allele | DRB3*01:01 in combination with another DRB3 allele | DRB3 negative | DRB3 allele other than *01:01 | No alleles associated with DRB3 | Only 1 allele associated with DRB3 | Two alleles associated with DRB3 | Two alleles associated with DRB3*01:01 | DRB3*01:01 negative | One dose of DRB3*01:01 | Two doses of DRB3*01:01 | |
| Only DRB3*01:01 | DRB3 allele other than *01:01 | |||||||||||
| 7 | — | — | — | 7 | — | — | — | — | — | 7 | — | — |
| — | — | 2 | — | — | — | — | — | — | — | 2 | — | — |
| — | — | — | 19 | — | — | — | — | — | — | — | 19 | — |
| — | 87 | — | — | — | — | — | 87 | — | — | — | 87 | — |
| — | 13 | — | — | — | — | — | — | 13 | 13 | — | — | 13 |
| — | — | — | — | — | — | 1 | — | — | — | 1 | — | — |
| — | — | — | — | — | 1 | — | — | — | — | 1 | — | — |
Typing was performed at the Tissue Typing Laboratory, Department of Immunology, Rikshospitalet, Oslo University Hospital, Oslo, Norway, a laboratory that is accredited by the European Federation for Immunogenetics.
In 100 women, sequencing of HLA-DRB3 gene only revealed the *01:01 allele. In 87 of these women, first-field resolution HLA-DRB1 typing demonstrated that they only carried 1 HLA-DRB1 allele with known association to the HLA-DRB3 gene. These cases were interpreted as hemizygous for the HLA-DRB3*01:01 allele. Thirteen women carried 2 HLA-DRB1 alleles that are known to be associated with the HLA-DRB3 gene. As first-field resolution HLA-DRB1 typing could not resolve the HLA-DRB3*01:01 zygosity status for these patients, second-field resolution HLA-DRB1 typing was performed. In all 13 cases, the women carried 2 HLA-DRB1 alleles, which are known to be associated with the HLA-DRB3*01:01 allele, and for this reason, these women were interpreted as being homozygous for the HLA-DRB3*01:01 allele.
Sequencing was not performed for 2 women. One of them was HLA-DRB3 typed and found only to carry the HLA-DRB3*02:02 allele. The other woman was HLA-DRB1 typed and found to carry 2 HLA-DRB1 alleles not associated with the HLA-DRB3 gene. Therefore, these women were interpreted as HLA-DRB3*01:01 negative.
The global analysis (Figure 1) demonstrated that the HLA-DRB3*01:01 allele had a significant dose-dependent effect on the maternal anti-HPA-1a levels (global P value [Pglobal] = .0032). The median (range) anti-HPA-1a levels were 1.5 IU/mL (0.0-19.0 IU/mL), 21.1 IU/mL (0.0-1967 IU/mL), and 43.7 IU/mL (1.0-980 IU/mL) in women with 0, 1, and 2 copies of HLA-DRB3*01:01 allele, respectively. The differences between 0 and 1 copies, as well as 1 and 2 copies, were also significant (P = .0063 and P = .0234, respectively).
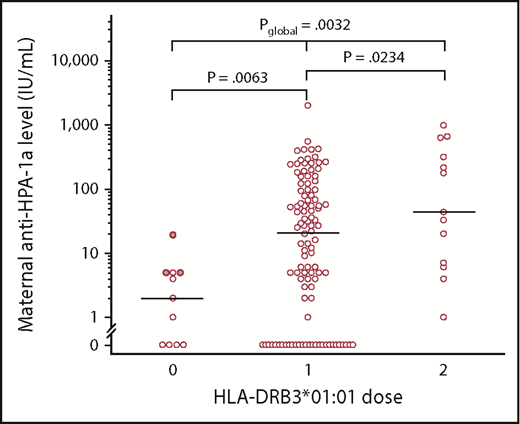
Maternal anti-HPA-1a levels (IU/mL) in relation to dose of the HLA-DRB3*01:01 allele. Antibody levels from HLA-DRB3*01:01–negative women, who carried the HLA-DRB3*02:02 allele, are marked with filled circles. Women who were heterozygous or hemizygous (1 dose) had higher antibody levels than HPA-1a–immunized HLA-DRB3*01:01–negative women (0 dose), but they had lower antibody levels than women who were HLA-DRB3*01:01 homozygous (2 doses). The horizontal bars denote the median values in each group of women. Kruskal-Wallis test was used to calculate Pglobal. The other P values were calculated by using the Mann-Whitney U test.
Maternal anti-HPA-1a levels (IU/mL) in relation to dose of the HLA-DRB3*01:01 allele. Antibody levels from HLA-DRB3*01:01–negative women, who carried the HLA-DRB3*02:02 allele, are marked with filled circles. Women who were heterozygous or hemizygous (1 dose) had higher antibody levels than HPA-1a–immunized HLA-DRB3*01:01–negative women (0 dose), but they had lower antibody levels than women who were HLA-DRB3*01:01 homozygous (2 doses). The horizontal bars denote the median values in each group of women. Kruskal-Wallis test was used to calculate Pglobal. The other P values were calculated by using the Mann-Whitney U test.
The opposite trend was noticed for neonatal platelet count (Figure 2), where the global analysis showed a significant dose-dependent effect of the mother’s HLA-DRB3*01:01 allele on the platelet count of the newborn (Pglobal = .0155). The median (range) neonatal platelet counts were 241 × 109/L (59 × 109/L to 393 × 109/L), 107 × 109/L (4 × 109/L to 387 × 109/L), and 32 × 109/L (4 × 109/L to 352 × 109/L) for newborns of women with 0, 1, and 2 copies of the HLA-DRB3*01:01 allele, respectively. Although the trend was significant, the difference in platelet count in neonates born of women with 1 copy of the HLA-DRB3*01:01 allele vs neonates born of women with 2 copies of this allele (Figure 2) did not reach significance (P = .0892).

Neonatal platelet count (×109/L) in relation to the mother’s dose of the HLA-DRB3*01:01 allele. Platelet counts of children born of HLA-DRB3*01:01–negative women, who carried the HLA-DRB3*02:02 allele, are marked with filled circles. Women who were heterozygous or hemizygous (1 dose) gave birth to newborns with lower platelet counts than children born of HPA-1a–immunized HLA-DRB3*01:01–negative women (0 dose), but the neonatal platelet count was higher than in children born of women who were HLA-DRB3*01:01 homozygous (2 doses). The horizontal bars denote the median values in each group of women. Values below the dotted lines, equivalent to platelet counts of 50 and 30 × 109/L, respectively, indicate particularly high risk of ICH. Of the children born of the 13 HLA-DRB3*01:01 homozygous women, 8 and 6 had platelet counts below these limits. For the 106 women with 1 dose of HLA-DRB3*01:01, these figures were 28 and 38, respectively. All the children born of women who lacked HLA-DRB3*01:01 had platelet counts >50 × 109/L. Kruskal-Wallis test was used to calculate Pglobal. The other P values were calculated by using the Mann-Whitney U test.
Neonatal platelet count (×109/L) in relation to the mother’s dose of the HLA-DRB3*01:01 allele. Platelet counts of children born of HLA-DRB3*01:01–negative women, who carried the HLA-DRB3*02:02 allele, are marked with filled circles. Women who were heterozygous or hemizygous (1 dose) gave birth to newborns with lower platelet counts than children born of HPA-1a–immunized HLA-DRB3*01:01–negative women (0 dose), but the neonatal platelet count was higher than in children born of women who were HLA-DRB3*01:01 homozygous (2 doses). The horizontal bars denote the median values in each group of women. Values below the dotted lines, equivalent to platelet counts of 50 and 30 × 109/L, respectively, indicate particularly high risk of ICH. Of the children born of the 13 HLA-DRB3*01:01 homozygous women, 8 and 6 had platelet counts below these limits. For the 106 women with 1 dose of HLA-DRB3*01:01, these figures were 28 and 38, respectively. All the children born of women who lacked HLA-DRB3*01:01 had platelet counts >50 × 109/L. Kruskal-Wallis test was used to calculate Pglobal. The other P values were calculated by using the Mann-Whitney U test.
Four of the 13 women who were homozygous for the HLA-DRB3*01:01 allele had rather low HPA-1a antibody levels (1, 4, 6, and 7 IU/mL, respectively). Three of these gave birth to children with normal platelet counts, whereas the fourth had a mildly thrombocytopenic child (platelet count, 130 × 109/L). The other 9 women had antibody levels ≥20 IU/mL; 8 of them gave birth to severely thrombocytopenic children (platelet count, ≤33 × 109/L).
In 1 case, fetal ICH was suspected after ultrasonographic examination at gestational week 34. The anti-HPA-1a level of the mother, who was heterozygous for the HLA-DRB3*01:01 allele, was 411 IU/mL. She was treated with corticosteroids for fetal lung maturation, and the next day, a girl (weight, 2320 g; Apgar scores, 9 and 10 at 1 and 5 minutes, respectively) was delivered by Caesarean section. The platelet count immediately after birth was 13 × 109/L; the child had widespread petechiae and was therefore transfused with platelets from an HPA-1a–negative donor. Subsequent magnetic resonance brain imaging revealed a large bleed (6 × 3 × 5 cm in size) in the left temporal region. ICH was not found in any other fetuses or newborns.
Discussion
The current study is the first to examine whether there is a dose effect of the HLA-DRB3*01:01 allele on maternal anti-HPA-1a levels and neonatal platelet counts in a cohort of prospectively identified HPA-1a–immunized women. The results show that the HLA-DRB3*01:01 allele imposes a clear dose effect on the level of maternal anti-HPA-1a levels, with an opposite trend for neonatal platelet counts. These results are in accordance with the general notion that development of anti-HPA-1a requires help from HPA-1a–specific T cells. Crystallographic studies have demonstrated that the peptide from the human platelet integrin β3 glycoprotein, which harbors leucine at residue 33 (the HPA-1a epitope), fits perfect into the cleft of the HLA molecule that is encoded by HLA-DRA/DRB3*01:01,21 whereas this peptide does not fit into the HLA molecules encoded by other HLA-DRB3 alleles, such as HLA-DRB3*02:02. Furthermore, CD4+ T cells, which specifically recognize integrin β3 peptides that include leucine at residue 33, have been isolated from HPA-1a–immunized women who have given birth to children affected by FNAIT.22-24 These T cells exclusively recognized the antigenic peptide when presented in the context of the HLA molecule encoded by HLA-DRA/HLA-DRB3*01:01.22-24
The impact of dose of the restricting element has also been demonstrated in celiac disease, where an inflammatory T cell response to HLA-DQ2–bound gluten peptides causes the disease. Vader et al25 have demonstrated that the HLA-DQ2 gene dose has a strong quantitative effect on the magnitude of gluten-specific T-cell responses in terms of T-cell proliferation and cytokine secretion (interferon-γ, tumor necrosis factor α, interleukin-10 [IL-10], IL-5, and IL-4) and that this correlates with the level of (gluten) peptide binding to HLA-DQ2 homozygous and heterozygous antigen-presenting cells (APCs).
When translating this knowledge to HPA-1a immunization, it is likely that APCs with double dose of the HLA molecule encoded by HLA-DRA/HLA-DRB3*01.01 (ie, HLA-DRB3*01:01 homozygous) bind more of the antigenic peptide and consequently provide a more efficient activation of the HPA-1a–specific T cells than APCs from HLA-DRB3*01:01 hemizygous or heterozygous women. This amplified T-cell response could provide more efficient help to HPA-1a–specific B cells, which differentiate into anti-HPA-1a–producing plasma cells, resulting in higher antibody levels in HLA-DRB3*01:01 homozygous women than hemi- or heterozygous women.
Wienzek-Lischka et al11 recently examined whether HLA-DRB3*01:01 zygosity status has any impact on neonatal platelet counts and found that the platelet count was independent of whether the mother carries 1 or 2 doses of the HLA-DRB3*01:01 allele. The population they examined originated from mother-child samples that were referred to their reference laboratory due to suspicion of FNAIT. In their study, the median neonatal platelet counts were ∼15 × 109/L and 20 × 109/L for women with 1 and 2 copies of the HLA-DRB3*01:01 allele, respectively.11 This is in clear contrast with our observation of much higher median platelet counts in children born of heterozygous women (107 × 109/L) as compared with children born of homozygous women (32 × 109/L). Hence, the very low platelet counts in the study by Wienzek-Lischka et al clearly indicate that there has been a selection bias toward more severe cases. Consequently, this bias may have prevented Wienzek-Lischka et al11 from detecting an effect of HLA-DRB3*01:01 zygosity status on neonatal platelet counts.
We also found the lowest platelet counts in children born of HLA-DRB3*01:01 homozygous women, followed by children born of women who were HLA-DRB3*01:01 hemizygous or heterozygous and finally children of HLA-DRB3*01:01–negative women. This finding is in accordance with a recent systematic review in which it was shown that there is a significant association between neonatal platelet counts and maternal anti-HPA-1a levels.26 The dose effect of the HLA-DRB3*01:01 allele on neonatal platelet count was not as obvious as the effect on anti-HPA-1a levels in these 3 groups of women. This is not surprising, because the causal chain of events between the maternal HLA-DRB3*01:01 allele dose and the mother’s anti-HPA-1a level is shorter than the chain of events between maternal HLA dose and neonatal platelet count. In addition to the level of anti-HPA-1a, the neonatal platelet count depends on a number of other factors, such as (1) the rate of transplacental transport of anti-HPA-1a; (2) the density of HPA-1a on fetal platelets; (3) the glycosylation pattern of the Fc-part of anti-HPA-1a IgG, which may regulate anti-HPA-1a’s affinity to the Fc receptor on the splenic macrophages27 ; and (4) the extent to which fetal thrombopoiesis can keep up with peripheral destruction of anti-HPA-1a–sensitized platelets.
The zygosity status of many of the women were inferred from the well-known linkage disequilibrium between the HLA-DRB1 and HLA-DRB3 genes, but rare associations of HLA-DRB1 and HLA-DRB3 genes have been described. Hence, in a population of 123 555 donors from the National Marrow Donor Program registry, unusual HLA-DRB1 and HLA-DRB3 associations were identified in 5 donors.28 However, as such associations are extremely rare, it is not likely that this could have biased the general result of the current study.
As several of the samples had anti-HPA-1a levels above the internal standard of 15 IU/mL, it was necessary to dilute these samples in order to obtain an antibody measurement within the linear part of the standard curve. However, we believe that the potential inaccuracy this may have introduced is of minor importance, primarily because the quantitative MAIPA assay with this particular internal standard has been validated according to guidelines from the International Conference on Harmonization.29 The robustness of this assay has also been documented in 2 interlaboratory studies.18,30
There is an unmet medical need of better tools for risk assessment of pregnant women who are HPA-1a immunized. The assessment of whether an HPA-1a–negative mother carries 1 or 2 copies of the HLA-DRB3*01:01 allele could potentially serve as tool a for clinicians in order to determine the level of clinical follow-up and treatment during pregnancy (eg, how often women should be monitored during pregnancy). However, as approximately one-third of women who are hemi- or heterozygous for the HLA-DRB3*01:01 allele give birth to severely thrombocytopenic children (Figure 2), it is questionably to what extent testing for zygosity status alone would provide guidance for the clinicians. It is possible that such an approach could one day complement future diagnostic tools to provide a better risk assessment for HPA-1a–immunized women.
Acknowledgments
The authors are grateful to Kirsten Sørensen (Blood Bank of Oslo, Oslo University Hospital) for sequencing exon 2 of the HLA-DRB3 gene of 128 of the included women, and Anne Kari Tveter (Tissue Typing Laboratory, Oslo University Hospital) for performing HLA-DRB3 and HLA-DRB1 typing of selected samples.
This study was primarily funded by internal research grants at Oslo University Hospital in addition to a grant from the European Union’s 7th Framework Program through the PROFNAIT project (grant agreement 305986).
Authorship
Contribution: J.K.-K. conceptualized the study, performed the final statistical analyses, and wrote the first draft of the manuscript; T.L.T. and M.K. gathered the data; T.L.T. made the first series of statistical analyses; B.A.L. and J.T.V. were responsible for the supplementary HLA typing and provided academic input related to interpretation of the HLA typing results; M.K. provided academic input regarding FNAIT; and all authors revised the manuscript and approved the final version.
Conflict-of-interest disclosure: J.K.-K. and M.K. belong to the group of founders and owners of Prophylix AS, a Norwegian biotech company that leads the European Union–founded PROFNAIT Consortium, which is developing a hyperimmune anti-HPA-1a IgG for the prevention of fetal and neonatal alloimmune thrombocytopenia. The remaining authors declare no competing financial interests.
Correspondence: Jens Kjeldsen-Kragh, Department of Clinical Immunology and Transfusion Medicine, University and Regional Laboratories, Region Skåne, Akutgatan 8, S-221 85 Lund, Sweden; e-mail: jkk@prophylix.com.

